MERISTEM CULTURE OF MINIATURE MONOPODIAL ORCHIDS
|
|
|
- Todd Cobb
- 5 years ago
- Views:
Transcription
1 MERISTEM CULTURE OF MINIATURE MONOPODIAL ORCHIDS A THESIS SUBMITTED TO THE GRADUATE DIVISION OF THE UNIVERSITY OF HAWAII IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF SCIENCE IN HORTICULTURE MAY 1972 By Oradee Intuwong Thesis Committee: Yoneo Sagawa, Chairman Haruyuki Kamemoto Douglas J, C. Friend

2 We certify that we have read this thesis and that in our opinion it is satisfactory in scope and quality as a thesis for the degree of Master of Science in Horticulture, THESIS COMMITTEE / Chg4.rman


3 TABLE OF CONTENTS Page LIST OF TABLES... iv LIST OF FIGURES... ACKNOWLEDGEMENT... v vi INTRODUCTION... 1 REVIEW OF LITERATURE... 3 MATERIALS AND METHODS... 8 RESULTS DISCUSSION SUMMARY LITERATURE CITED... 65

4 LIST OF TABLES Page TABLE 1. THE ABILITY OF DIFFERENT HYBRIDS AND BUDS FROM DIFFERENT POSITIONS TO FORM PROTOCORM-LIKE BODIES TABLE II. PROLIFERATION OF TISSUE FROM YOUNG INFLORESCENCES... 15


5 LIST OF FIGUEES Page FIGUEE 1. METHOD OF OBTAINING EXPLANTS FIGUEE 2. INFLOEESCENCE CULTUBE FIGURE 3. BUDS GEOWN IN SOLID VAGIN AND WENT FOR 1-2 MONTHS FIGUEE 4. MORPHOLOGICAL ORIGIN OF PEOTOCORM-LIKE BODIES FIGURE 5. PATTERN OF EXPLANT DEVELOPMENT FIGUEE FIGURE FIGURE FIGURE 6. GROWTH OF PROTOCOEM-LIKE BODIES FROM TERMINAL AND AXILLARY BUDS IN AGITATED LIQUID MEDIUM GROWTH OF PROTOCOEM-LIKE BODIES IN AGITATED LIQUID MEDIA PLANTLETS FROM PROTOCOEM-LIKE BODIES GROWN IN AGITATED LIQUID MEDIUM ANATOMY OF PROTOCOEM-LIKE BODIES WITH MERISTEMATIC AREAS FIGURE 10. ANATOMY OF MERISTEMATIC AREA WITH LEAF PRIMORDIA 36 FIGURE 11. LONGITUDINAL AND CROSS SECTIONS OF PROTOCOEM-LIKE BODIES FIGURE 12. LONGITUDINAL SECTION OF YOUNG INFLORESCENCE FIGURE 13. CROSS SECTION OF YOUNG INFLORESCENCE FIGURE 14. CROSS SECTION OF OLDER PORTION OF INFLORESCENCE 45 FIGURE 15. CULTURES FROM YOUNG INFLORESCENCES FIGURE 16. GROWTH OF PROTOCORM-LIKE BODIES FROM YOUNG INFLORESCENCES IN AGITATED LIQUID MEDIUM FIGURE 17. ONE-LEAF STAGE PROTOCORM-LIKE BODIES FIGURE 18. GROWTH OF PROTOCORM-LIKE BODIES AND PLANTLETS ON DIFFERENT MEDIA FIGURE 19. GROWTH OF PROTOCORM-LIKE BODIES ON SOLID MEDIA... 57
6 ACKNOWLEDGEMENT The author expresses deep appreciation to the East-West Center for the scholarship to pursue graduate work at the University of Hawaii, the American Orchid Society for partial financial support for equipment, and to Christopher Teo for technical assistance in writing.
.")

7 INTRODUCTION The shoot-tip (meristem) culture technique has been widely used for the clonal propagation of orchids. This method is probably the biggest breakthrough in orchid propagation since the early 1920s, when the method of asymbiotic seed germination was reported by Knudson (1922). Propagation by shoot-tip culture makes possible the production of numerous identical plants from commercially desirable hybrids which are generally extremely heterozygous within a short period of time. Current asexual methods of propagation such as division of rhizomes or topping of the mother plant yield only a limited number of plants. Thus, they are slow and costly. Meristem culture is now being used on a commercial scale with Cymbidium (Morel, 1960, 1964; Sagawa _et al,, 1966), and Cattleya (Reinert and Mohr, 1967; Scully, 1967; Lindemann, 1970), Other sympodial orchids which have been successfully propagated by meristem culture include Dendrobium, Calanthe, Odontonia, Vuylstekeara, Spathoglottis, Crytopodium, Lycaste, Miltonia, Odontoglossum, Phaius, Paphiopedilum, Oncidium, and Zygopetalum (Morel, 1965, 1971; Vaucherot, 1966; Bertsch, 1967; Sagawa et al,, 1967), The monopodial type of orchid like Phalaenopsis and Vanda does not or barely responds to the standard techniques (Scully, 1967), Vajrabhaya (1970) reported only limited success of Rhynchostylis gigantea in meristem culture. The main purpose of this study was to find out whether miniature monopodial orchids could be propagated by meristem culture as successfully as sympodial orchids. In addition, if they did respond to
8 2 meristem culture, the following were sought: (1) to determine what sources of explants could most readily be induced to form protocorm-like bodies (plbs); (2) to study the growth pattern, and the effect of agitation on the differentiation of protocorm-like bodies into plantlets; (3) to find a suitable medium for proliferation, differentiation and growth of tissues up to the plantlet stage*


9 LITERATURE REVIEW Since Morel (1960) demonstrated the usefulness of meristem (shoot- tip) culture as an asexual method of propagation as well as of eliminating virus from orchids, this technique has been discussed extensively among orchid growers. The various techniques for successful culture of shoot and leaf tips were described by various workers (Morel, 1960, 1964; Sagawa et al., 1966; Reinert and Mohr, 1967; Lindemann, 1970; Churchill t, 1970, 1971). In the sympodial orchids like Cattleya, Cymbidium, Dendrobium, a young shoot arising from a "back bulb" or from a "green bulb" provided suitable material for a source of explants. Kim ^ l. (1970) showed that the best source of explant in Dendrobium came from those shoots weighing between 1-3 gm and having an average length of 5 cm. Explants from apical buds were most likely to form plbs. Vajrabhaya (1970) used axillary and terminal buds. Personal communication with Iwanaga and Taba Lab (2610 Waiomoa Rd., Honolulu 96816) indicated that newly initiated flower stalks (0.75 cm) can be used as a source of explants. Wimber (1965) recognized that leaf tissue of some aseptically grown Cymbidium plantlets in liquid medium produced plbs when constantly buffeted against the sides of the glass flasks at 200 rpm. Churchill _et al. (1970, 1971) were able to induce callus production from the leaf tips of seedlings and mature Cattleya, Laeliocattleya and Epidendrum plants. Only the young tissues obtained before the development of the notch so typical of Cattleya and Laeliocattleya leaf tips could form callus. Tissues removed after the notch was
, Similarly, Reinert and Mohr (1967) showed that the growth of Cattleya explants was generally")
10 4 evident did not produce callus. Attempts to induce callus from other parts of the leaf failed. Liquid vs. Solid Media and Effect of Agitation Tissues grow better in liquid cultures than on agar (Steward 1952), Similarly, Reinert and Mohr (1967) showed that the growth of Cattleya explants was generally faster and callus development was more frequent in liquid medium than on agar. When placed directly on an agar medium during the first three weeks of growth, excised meristem tissues turned brown and subsequently died, whereas, tissues submerged in liquid culture grew and developed normally. Apparently the tissues receive enough oxygen through agitation of the culture. For Cymbidium and Dendrobium solid or liquid media served equally well (Sagawa and Kunisaki, 1968), Wimber (1963) found that in a flask which was not agitated, the meristem tissue followed a developmental sequence which superficially resembled that of seed germination, i,e,, after the protocorm had reached about 4 mm size, a shoot appeared. In flasks which were agitated, these plbs did not generally produce shoots but continued to proliferate into more plbs. It was considered that shaking apparently checked any polarity that might develop in the tissue and this inhibited shoot formation, possible effects of agitation; Scully (1967) listed the following (1) to dilute some specific inhibitor produced in response to excision and sterilization^ (2) to improve aeration,.ultimately providing for full activity in respiration, protein synthesis, and salt uptake; (3) or, in case of proliferating
.")
11 tissues, to increase the surface area exposed to nutritive materials. 5 Nutrient Media The requirements for tissues in culture are little different from the requirements of germinating seeds (Wimber, 1963; Morel, 1965). Simple inorganic nutrient media with sugar and coconut water were the most suitable general culture medium (Sagawa and Kunisaki, 1968). Morel (1965) found that the complex media containing plant extracts like banana, tomato juice or coconut water used for seedlings gave poor results, and Knudson C medium was preferred. There were, however, some workers (Lindemann, 1967; Vajrabhaya, 1970; Churchill et al,, 1970, 1971) who showed that certain species of orchids needed more than just simple media in order for tissues to proliferate. Vajrabhaya (1970) formulated a medium for culturing tissue of Rhynchostylis, a monopodial orchid, which responded poorly to standard techniques. Lindemann (1967) found that using "starting, maintenance and rooting" media for Cattleya culture gave better results than just simple, uniform nutrient media used throughout the stage from proliferation to plantlet. In culturing leaf tips, Churchill ejt al_, (1970, 1971) used modified Murashige-Skoog and modified Heller's medium to obtain callus. To obtain plantlets from the callus and protocorms produced, Knudson C medium was used. The mineral requirements of a given tissue is not fixed but depends on its physiological state. The biochemical-physiological differentiation in orchids appeared to parallel the more evident morphological-anatomical differentiation (Arditti, 1965), Thus, in

12 principle, the chemical composition of nutrient solution should be adapted to each particular growth stage, and even the size of tissue at a particular time. Requirements for growth factors vary according to the species, and even to the nature of the tissues from one and the same species. Most plant tissues are not carbon autotrophic. Even if the initial explant is rich in chlorophyll, it becomes more or less completely depigmented during the culture, and photosynthesis ceases. The tissues must then be cultivated in sugar-containing media (Ueda and Torikata, 1968). Gautheret (1955) showed that sucrose was the best carbon source followed by glucose, maltose, raffinose, fructose and galactose. Galactose caused damage and retarded the growth of seedlings (Ernest et ^., 1971). Pentoses and polysaccharides cannot be utilized by normal carrot tissue. Sucrose had been used in concentrations ranging from for good growth, Yates and Curtis (1949) grew Cattleya on media containing 0.06 to 0,24 molar sucrose. As the plants became older (about one year old), increased root growth was conspicuous and shoot growth was notably.decreased with higher concentration of sucrose. Ueda and Torikata (1968) showed that increasing the sucrose concentration of the media considerably increased the fresh weight and branching of Gymbidium protocorms, the most abundant growth being observed with 47. sucrose. According to Hayes (1969), although sugar was necessary for practical success, its requirement was not absolute, Arditti (1965) stated that sugar free media were incapable of supporting any appreciable growth regardless of additives.
.")
13 The use of coconut water for ^ vitro culture was first proposed by van Overbeek t al«in 1941, The most effective concentrations vary from 5 to 257o, Addition of coconut water induced strong proliferation but retarded differentiation of Phalaenopsis seedlings (Ernest, 1967). When provided in the germination medium, coconut water inhibited the growth of Dendrobium seedlings but stimulation of growth was observed in one year old seedlings (Kotomori and Murashige, 1965), Control of Differentiation Root and shoot initiation from solid cultures (Murashige, 1965) and agitated liquid cultures (Bonnett and Torrey, 1965) could be controlled to some extent by changing the type of growth regulator used (Maheshwari and Gupta, 1965; Kent and Steward, 1965) or altering its concentration (Staba et_ al^., 1965), Coconut water (Halperin, 1966bj Steward e^ al^,, 1964) and reduced nitrogen (Reinert et al,, 1966; Halperin, 1966a) may or may not be needed to induce plantlets. The technique of inducing plantlets is sufficiently advanced so that thousands of similar Cymbidium plants were produced within a year from a meristem (Morel, 1964). The ability to regulate plbs so that they grow continuously, as either undifferentiated tissues (plbs) or as differentiated tissues (plantlets), is of scientific as well as commercial significance.


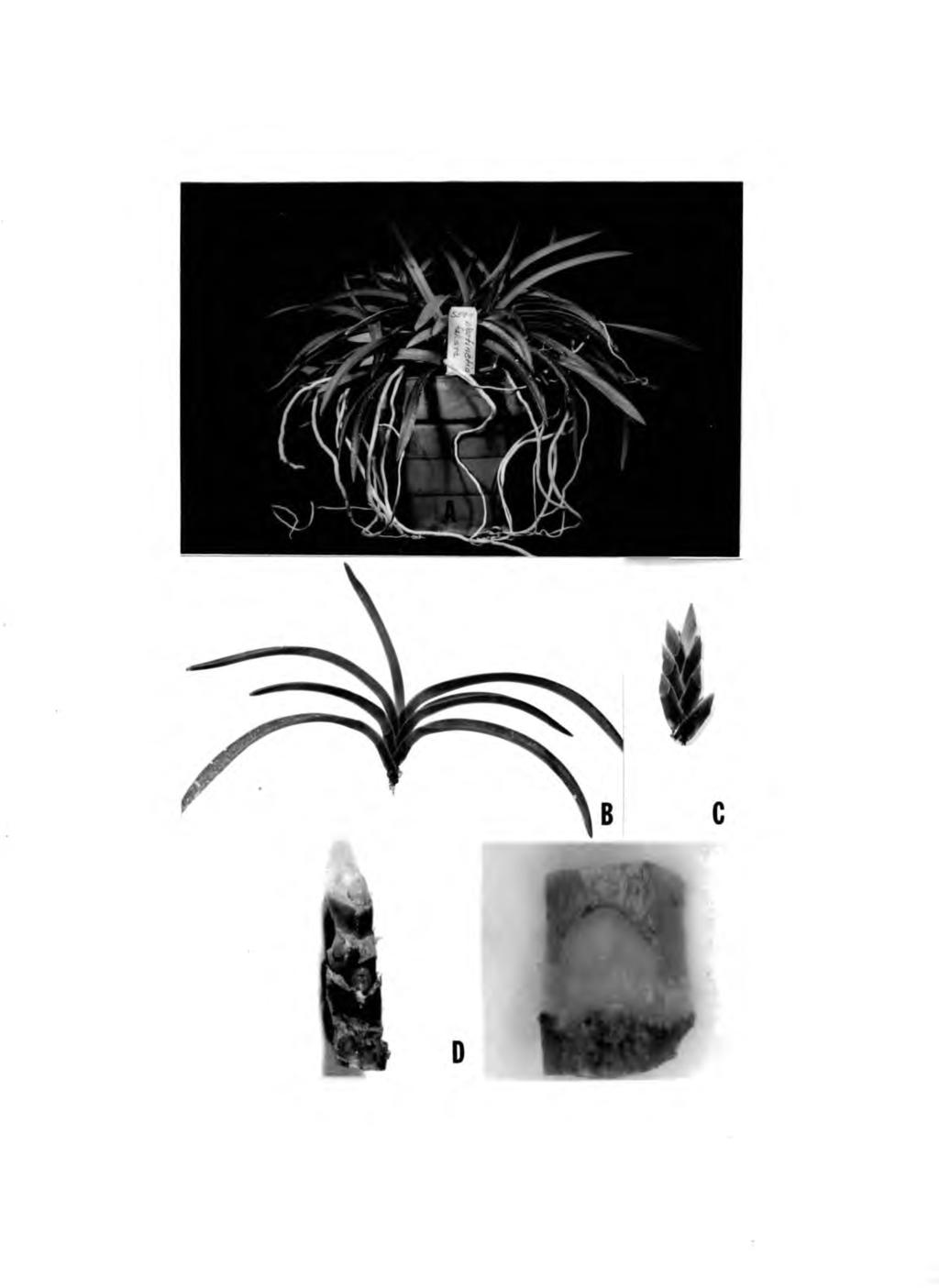
14 MATERIALS AND METHODS The miniature monopodial orchids used in this study were obtained from the collection of the Horticulture Department, University of Hawaii. The following hybrids were used: Name Hybrid code number Parents Ascofinetia Twinkle Ascofinetia Twinkle Ascofinetia Cherry Blossum Vandofinetia Premier Neostylis Lou Sneary Neostylis Lou Sneary Vascostylis Blue Fairy Neofinetia falcata x Ascocenda miniatum Neofinetia falcata x Ascocenda miniatxjm Neofinetia falcata x Ascocenda ampullaceum Neofinetia falcata x Vanda lamellata Neofinetia falcata x Rhynchostylis coelestis Neofinetia falcata x Rhynchostylis coelestis Ascocenda Meda Arold x Rhynchostylis coelestis Sources of explants used were from vegetative shoots of the six to seven leaf stage (Figs.lA, B). The leaves of the shoots were removed (Fig. 1C) and the shoot axis was then sterilized for 15 minutes in 10% clorox with one drop of Tween-20 in 100 ml liquid. The leaf-bases still attached to the shoot axis were removed (Fig. ID) The shoot axis was again placed in 5% clorox solution for 10 minutes. To remove traces of clorox, the sterilized.materials'were then soaked in sterile distilled water for three minutes.
. Shoot axis after leaves were removed (C X 1/2).")
15 Fig. 1. Method of obtaining explants. Plants used for meristem culture (A x 1/4). Vegetative shoot of seven-leaf stage, a suitable source of explant (B x 1/3). Shoot axis after leaves were removed (C X 1/2). Axis of stem with terminal and axillary buds (D x 1). An explant from an axillary bud (E x 10).

16 E 10
, The size of the buds was approximately 2-3 mm (Fig, IE), The buds in the detached shoots were numbered basipetally, (2) the terminal buds,")
17 11 Two kinds of buds were used as explants: which are located in the axil of the leaves. (1) the axillary buds, Usually three to seven buds were found on a stem. They are almost entirely covered by flat scales (Fig, ID), The size of the buds was approximately 2-3 mm (Fig, IE), The buds in the detached shoots were numbered basipetally, (2) the terminal buds, which consisted of the apical meristem plus two to four leaf primordia. Both axillary and terminal buds were cultured in vials (4" x 1") on Vacin and Went medium (1949) modified by the addition of 157o coconut water (v/v). For solid medium 97. agar (w/v) was added. Cultures were placed under 200 foot-candle continuous illumination, obtained from G.E. white fluorescent lamps (Power Groove). The temperature was C. After one or two months of culture on agar medium, the scale-like leaves which enclosed the growing tissue were removed and the tissues were transferred into liquid medium. The 50 ml erlenmeyer flask containing 20 ml of medium was continuously agitated at approximately 160 rpm using a New Brunswick Model V shaker. After four to five weeks in the agitated medium, plbs began to form. In some cases, removal of scale-like leaves did not result in. proliferation and scale leaves developed instead. These needed to be removed again to induce proliferation. Inflorescences with floral primordia at various stages of differentiation were removed from the shoots and measured for length for Inflorescence culture. Sterilization was done as described earlier. After the bracts were removed, these inflorescences were cultured in

18 12 agitated liquid medium. Once plbs were obtained, the amount of sucrose in the medium was reduced to zero. Otherwise the plbs turned yellow. Plbs were subcultured into fresh liquid medium every two to three weeks. For anatomical study, tissues were killed and fixed in Graf fixative, dehydrated in a series of tertiary bytyl alcohol, and embedded in paraffin. Sections cut 10 to 12j a ~ thick were stained by the rapid Safranin and Fast Green technique (Shapiro, 1947). For morphological and anatomical observations and photography, a Wild M-5 stereo-microscope or Zeiss photomicroscope was used.
 of these were contaminated with fungus or bacteria. Out of the 83 uncontaminated cultures, 26 (31.37,) proliferated within 58-118 days and produced plbs.")
19 RESULTS 1. Study on the ability of different hybrids and buds from different positions to form plbs A total of 136 explants were cultured and 53 (39.07,) of these were contaminated with fungus or bacteria. Out of the 83 uncontaminated cultures, 26 (31.37,) proliferated within days and produced plbs. The percentage of cultures that formed plbs varied from depending on the hybrids (Table I). The highest percentage of success in plb formation was obtained with hybrid 1873, while the lowest was with hybrid As shown in Table I explants of apical buds resulted in plb formation. Axillary buds from positions one to four resulted in 33.3 to plb formation. The buds from positions five and below did not proliferate. In inflorescence culture, inflorescences of 1% cm or less proliferated (Fig. 2A) within 20 to 51 days (Table II), whereas those which were three cm or longer either had elongated internodes and florets which turned brown (Fig. 2B) or florets which developed and bloomed (Fig. 2C).
 1/1 (68) 3/3 (107) 0/1 0/1. 2/2 8/12 66.7 Axillary buds Position 1 0/2 1/3 (96) 1/4 (98) 2/7 (118) 1/4 (61) 3/4 (90) 8/24 33.")
20 14 TABLE I, The ability of different hybrids and buds from different positions to form protocorm-like bodies Sources Hybrid number io uai 11 ^ rerceni. Apical buds 2/4^ (58) 1/1 (68) 3/3 (107) 0/1 0/1. 2/2 8/ Axillary buds Position 1 0/2 1/3 (96) 1/4 (98) 2/7 (118) 1/4 (61) 3/4 (90) 8/ Position 2 0/1 0/0 1/4 (60) 0/3 1/4 (114) 2/4 (107) 4/ Position 3 0/1 2/2 (104) 0/1 1/6 (107) 0/3 1/3 (103) 4/ Position 4 0/0 0/0 0/1 1/6 (114) 1/3 (61) 0/0 2/ Position 5 0/0 0/0 0/0 0/1 0/1 0/0 0/2 0 Position 6 0/0 0/0 0/0 0/1 0/1 0/0 0/2 0 Position 7 0/0 0/0 0/0 0/0 0/1 0/0 0/1 0 Total 2/8 4/6 5/13 4/25 3/18 8/13 26/83 Perc.ent a b The axillary buds are numbered basipetally Nvtmbers of cultures forming plbs/total number of uncontaminated cultures ( ) Average number of days to proliferate


21 15 TABLE II. Proliferation of tissues from young inflorescences Hybrid number Inflorescence number Inflorescence length (cm) Proliferation Days to proliferate no proliferation + proliferation
.")
22 16 Fig. 2. Inflorescence culture. Proliferation of tissue from young inflorescence (A x 1.2). Unsuccessful culture of young inflorescence. Pedicels were elongated and florets turned brown and died (B x 1.2). Unsuccessful culture of young inflorescence in which florets developed and bloomed (C x 2.5).

23 17

, while several shoots developed from the terminal bud explant (Fig. 3B).")
24 18 2. Growth pattern of explants and differentiation into plantlets A. Explants from axillary and terminal buds The explants increased in size for one to two months in culture on solid VW medium without tissue proliferation. Scale-like leaves were visible surrounding the apical meristem. If these scale-like leaves were not removed from explants of axillary buds a shoot developed (Fig. 3A), while several shoots developed from the terminal bud explant (Fig. 3B). However, proliferation of tissues into plbs started four to five weeks after removal of these scale-like leaves and culture in agitated liquid medium (modified VW). Plbs were observed to originate from various points on the explant. (1) Tissues proliferated from the cut surface of the scale-like leaves at the nodal area (Fig. 4A). (2) Plbs were also formed from the surfaces of the young leaves (Fig. 4B). Usually proliferation was acropetal on the explant. Most of the initial explants which did not proliferate turned brown and degenerated slowly. The initially clear medium turned turbid to brown with time. If the medium was not changed after 10 to 14 days, growth of plbs slowed down. The pattern of explant development can be classified in one of the following three ways: (1) Explant grew directly into shoots without forming plbs (Fig. 5A). '

25 19 (2) Explant produced both one or more shoots and plbs (Fig. 5B). (3) Explant produced plbs only (Fig. 5C). Growth of plbs started slowly and increased rapidly with time (Fig. 6). Hybrid 1923 produced the highest fresh weight of plbs while hybrid 1873 the least. Plants within the same hybrid also differed in the fresh weight of plbs produced as illustrated by hybrids 1924 and Initially, plbs did not differentiate organs but kept on multiplying (Fig. 7A), several protuberances were formed and each green plb produced a number of similar bodies which remained attached to each other. However, leaves were visible 75 to 100 days after proliferation (Fig. 6). At this stage, the culture was still in agitated liquid medium (Fig. 7B). As more leaves began to develop the movement of plbs in the agitated medium became restricted due to crowding within the flask and, therefore, could be considered stationary (Fig. 7C). In hybrids 1923 and 1924, roots appeared 50 to 60 days after the tissue became stationary (Fig. 8A). Root hairs formed on roots of plbs that remained above the liquid medium surface (Fig. 8B). In hybrids 1908, 1907 and 1873, no roots were observed (Fig. 6). To encourage the growth of more plbs, fragments of plbs were cultured in agitated medium. As long as the tissues were subcultured every two weeks and flasks kept agitated and not congested, multiplication of plbs continued. A plb consisted of a mass of undifferentiated parenchyma cells. Several meristematic areas were observed within this mass (Fig. 9). These meristematic areas consisted of isodiametric cells which had
.")
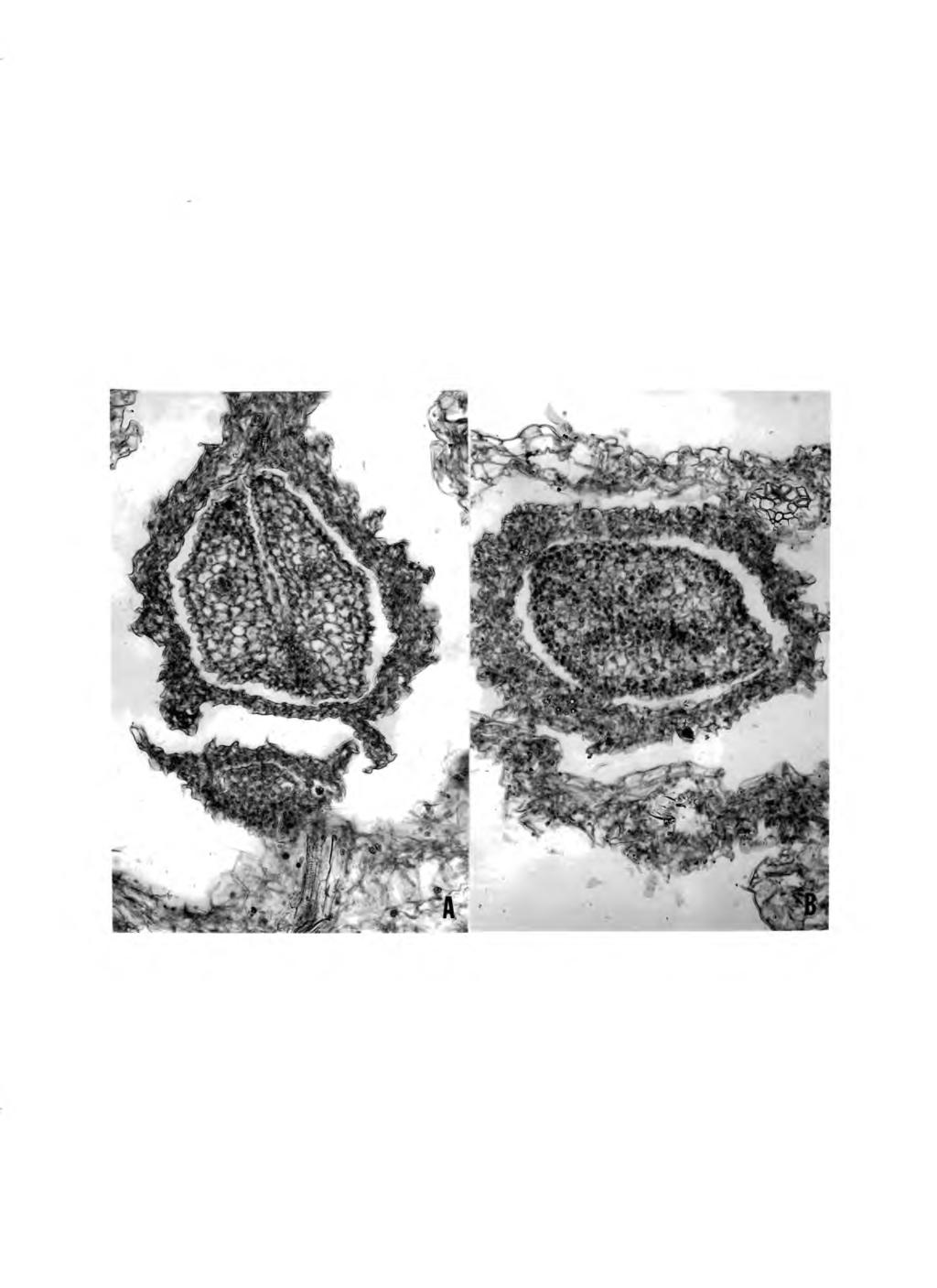
26 20 dense cytoplasm with prominent nuclei. From these cells originated hemispherical protuberances and leaf primordia (Fig. 10). No vascular system was observed in the plbs in the early stage prior to differentiation of leaves (Fig. 9). However, after leaf formation, paraffin sections of plbs revealed the presence of xylem elements (Fig. 11).
.")
27 21 Fig. 3. Buds grown in solid Vacin and Went for 1-2 months. Axillary bud from which a shoot developed (A x 15). Terminal bud from which several shoots developed (B x 15).

28 22
 and from")
29 23 Fig. 4. Morphological origin of protocorm-like bodies. Tissues proliferated at the cut surface of the scale-like leaves at the nodal area (A x 10) and from the surfaces of the young leaves (B X 10).
30 B 24
 (A x 10), explant produced")
31 25 Fig. 5. Pattern of explant development. Explant grew directly into shoots without forming protocorm-like bodies (plbs) (A x 10), explant produced both shoot and plbs (B x 10), and explant producing plbs only (C x 10).


32 B 26
33 27 Fig. 6. Growth of protocorm-like bodies (plbs) from terminal and axillary buds in agitated liquid medium. L = Leaves visible S= Stationary plbs in agitated media R = Roots visible

34 CD
35 29 Fig. 7. Growth of protocorm-like bodies (plbs) in agitated liquid media. The non-differentiated rapidly multiplying plbs (A x 1.2), plbs with differentiated leaves (B x 1.2), and plbs with differentiated leaves and roots (C x 1.2).


36 30 B C
, root hairs were formed on roots that remained on")
37 31 Fig. 8, Plantlets from protocorm-like bodies (plbs) grown in agitated liquid medium. Plbs differentiated into roots and leaves (A X 6), root hairs were formed on roots that remained on liquid surface (B x 6).

38 B 32
39 33 Fig. 9. Anatomy of protocorm-like bodies showing meristematic areas within the tissue mass (x 60).


40 34

41 35 Fig. 10. Anatomy of protocorm-like bodies showing leaf primordia derived from cells of the meristematic area (x 200).

42 36
 and cross sections (B) of protocorm-like")
43 37 Fig, 11. Longitudinal (A) and cross sections (B) of protocorm-like bodies showing the presence of xylem elements (x 200).

44 38

45 39 B. Explants from young inflorescence Paraffin sections 10 to 12yu were made from young inflorescences one week after proliferation (Figs. 12, 13). A layer of meristematic cells, derived from the epidermis, formed calli from which plbs developed. The parenchymatous cells of the cortex and pith did not show sign of meristematic activity, and numerous vascular bundles were developed. In the region where the bract was removed, no callus was formed. Fig. 14 showed that at the base of the proliferating plbs a necrotic layer was formed from the outer cortical cells of the original tissue. This necrotic layer caused the separation of plbs from the original explant. from the newly formed plbs. further and turned brown. Subsequent multiplication of plbs was The original tissues did not proliferate Tissues from the bracts (Fig. 15A) and florets (Fig. 15B) on the young inflorescence did not proliferate and eventually turned brown. The rate of growth of plbs from inflorescence culture is shown in Fig. 16. Initially growth was slow but after 50 days rapid growth ensued. Leaves appeared 86 days after proliferation. The flask began to get congested and within 20 days, the culture became stationary. Two months later, roots were formed.
46 40 Fig, 12. Longitudinal section of young inflorescence one week after proliferation showing layers of meristematic cells derived from the epidermis. The parenchymatous cells of the cortex and pith did not show sign of meristematic activity (x 60),

47 41
48 42 Fig. 13. Cross section of inflorescence one week after proliferation, showing meristematic cells formed on epidermis all over the surface of peduncle (x 60).

49 43
50 44 Fig. 14. Cross section of older portion of inflorescence, showing layers of necrotic cells which formed on the outer cortex of peduncle, caused the separation of protocorm-like bodies from the original explant (x 60).


51 45
 did not proliferate.")
52 46 Fig. 15. Young inflorescence culture showing tissue from bracts (A X 1.2) and florets (B x 1.2) did not proliferate. Protocorm-like bodies were formed from tissues at the base of inflorescence.

53 47
 from young inflorescence in agitated liquid")
54 48 Fig. 16, Growth of protocorm-like bodies (plbs) from young inflorescence in agitated liquid medium, L = Leaves visible S = Stationary plbs in agitating media R = Roots visible

55
 were transferred from the agitated liquid to the following solid media: (1) modified VW (2) modified VW without sucrose (3) VW (4) VW without sucrose The two-month old plb cultures are")
56 50 3. Study of various media suitable for plb growth into plantlets One-leaf stage plbs (Fig. 17) were transferred from the agitated liquid to the following solid media: (1) modified VW (2) modified VW without sucrose (3) VW (4) VW without sucrose The two-month old plb cultures are illustrated in Fig. 18A..Modified VW without sucrose medium resulted in the greatest multiplication of plbs and some plbs had two to more leaves. However, on this medium no roots were produced, but there were numerous rhizoids (Fig. 19A). On modified VW, plbs proliferated but eventually turned yellow to brown. On VW most of the plbs turned brown, although some grew into plantlets and produced roots (Fig. 19B). There was little proliferation and differentiation of plbs when grown on VW without sucrose. When cultures which were yellow were transferred to modified VW without sucrose, they became green and resumed growth. Two to three-leaf stage plbs from modified VW without sucrose medium were transferred to the four similar media list previously (Fig. 18b ). After three months plbs on modified VW turned brown. On modified VW without sucrose, most tissues proliferated into plbs, but only seven plantlets were obtained. Most plbs cultures on VW turned brown, but some plbs differentiated into plantlets. A total of 25 plantlets were obtained. Plbs grown on VW without sucrose were yellow with little proliferation and differentiation.
57 51 The plantlets from the V medium were grown on the same four different media listed earlier. On modified V, plantlets grew very rapidly and were 1% inches high three months later (Fig. 16C). Plantlets on VW without sucrose showed the slowest growth and some died three months later.
58 52 Fig. 17. One leaf-stage of protocorm-like bodies used for culture on solid medium (x 6),

59 53
60 54 Fig. 18. Growth of protocorm-like bodies and plantlets on four different media (left to right) Vacin and Went with coconut water and sucrose, Vacin and Went with coconut water without sucrose, Vacin and Went with sucrose, Vacin and Went with neither coconut water nor sucrose. Protocorm-like bodies transferred from modified liquid Vacin and Went without sucrose (A x 1/3). Two to three leaf- stage protocorm-like bodies transferred from modified solid Vacin and Went without sucrose (B x 1/3). Plantlets transferred from solid Vacin and Went with sucrose (C x 1/3).

61 55
62 56 Fig. 19. Growth of protocorm-like bodies (plbs) on solid media. Plbs on modified Vacin and Went without sucrose showed no roots but numerous rhizoids formed. Many plbs were produced (A x 5). Most plbs on Vacin and Went with sucrose turned brown but some plbs differentiated into plantlets (B x 5).

63 57
.")
 pointed out that only juvenile cells have the regenerative properties to form plbs, mature cells did not grow. As buds get older (i.e., position of buds get further from the apex), there was a decrease in the number of explants which form plbs.")
64 DISCUSSION 1. Study on the ability of different hybrids and different positions of explants to form plbs in culture The use of apical buds for culture resulted in a high percentage of culture forming plbs (66.7%). Axillary buds from position one to four gave lower success ( %) while buds from position five and below did not proliferate at all. Morel (1971) pointed out that only juvenile cells have the regenerative properties to form plbs, mature cells did not grow. As buds get older (i.e., position of buds get further from the apex), there was a decrease in the number of explants which form plbs. not suitable for culture. Buds below position five were therefore This observation is contradictory to the report that axillary buds in Dendrobium showed an acropetal decreased in success of culture (Kim ^ JLL* > 1970). Such difference may be due to the difference in growth habit of these two groups of plants. Dendrobium, sympodial orchid, may have less active buds acropetally, while the opposite is true of monopodial orchids. Plants of different hybrids or within the same progeny vary in their response to meristem culture. Hybrids 1873 and 1800 were both Ascofinetia Twinkle, but the ability of cultures to form plbs were % respectively. Neostylis Lou Sneary 1907 gave the lowest percentage of success. Such differences in success may be due to genetic variations of the plant material. It might be of interest to work at the species level to see where such variations originated. The use of floral buds and young inflorescence of dicotyledonous plants for culture have been reported by many workers (Konar and
, and inflorescence axis (Sehgal, 1968) proliferated and formed plantlets.")
65 59 Konar, 1966; Konar and Nataraja, 1969; Mohan Ram and Wadhi, 1968; Sehgal, 1968; and Ganapathy, 1969). The pedicel, torus, calyx (Konar and Nataraja, 1969), and inflorescence axis (Sehgal, 1968) proliferated and formed plantlets. Such proliferation was shown to originate from the epidermal cells all along the surface of the stem (Konar and Nataraja, 1969). This study showed that the young inflorescence of a monocotyledon could also be induced to proliferate and form plantlets. Similarly, the epidermal cells all along the surface of the peduncle (Figs, 12, 13) showed meristematic activity, formed calli and eventually developed into plbs. The use of young inflorescences for mericloning monopodial orchids showed great potential, and also had the following advantages over the use of terminal and axillary buds: (1) the procedure is relatively easy, (2) contamination of cultures is lower, (3) percentage of cultures forming plbs is higher, (4) plbs can be obtained sooner than those from terminal and axillary buds. This is because the explant was cultured directly in liquid medium, (5) this method does not require sacrificing a plant, and there is no setback of parent plant through removal of young inflorescences, 2. Growth pattern of explants and differentiation into plantlets The tissue from which plbs originate varies a great deal with genera and species. Morel (1971) reported that sometimes the outgrowth came from the central axis or leaf primordia as in Cymbidium, or
 as in the case of inflorescence culture.")
66 60 in Cattleya the axis remained inert and only the cells of the leaf epidermis proliferated. In the case of Vanda the cortex of the stem near the apex proliferated. In this study, the outgrowth came from the epidermis of the peduncle (Figs. 11, 12) as in the case of inflorescence culture. In the apical and axillary bud cultures, outgrowth originated from the cut surface of the scale-like leaves at the nodal area (Fig. 3A). Also in some cultures, young differentiated leaves proliferated (Fig. 3B). The time required for tissues to proliferate and form plbs varies depending on types of tissues and species. In this study, the axillary bud and terminal buds took two to three months (Table I) to proliferate, while within one month, plbs were formed from the young inflorescence culture (Table II). Other workers reported that Cymbidium took one to two months (Morel, 1965) to produce plbs, while Cattleya and Dendrobium (Sagawa _et al^., 1966, 1967) took one and a half months. The plbs formed from these cultures were very small compared to plbs from Cymbidium, Dendrobium and Cattleya* They did not form big clumps but separated by themselves while still on the shaker or on agar medium. For multiplication of plbs, no sectioning was necessary. In case of Cymbidivim, Dendrobium and Cattleya, to encourage multiplication, the plbs have to be fractionated (Wimber, 1965). Agitation of mediimi did not totally inhibit leaf differentiation of plbs (Fig. 6B). Leaves formed on plbs days (Figs. 5, 15) after proliferation. As the flask became congested with the growing plbs, movement of tissue stopped. Two months after this stage, roots

67 61 appeared. From this, it is suggested that agitation of the medium affected root but not leaf differentiation. As long as plbs were agitated, roots did not form. If multiplication of plbs is desired, plb culture needs to be kept constantly agitated. Wimber (1963) reported that if cultures of Cymbidium were kept agitated, shoot formation was inhibited and only plbs formed. In miniature monopodial orchids studied, this was not observed; leaf differentiation occurred in agitated medium. 3. Media suitable for plbs growth into plantlets Plbs sucrose. proliferated well on media with coconut water but without The presence of coconut water caused proliferation of tissues from the original explant, into callus, and also multiplication of plbs. Radley (1958) and Murashige (1961) suggested that gibber- rellin, found in coconut water, inhibited organ differentiation. O However, Reinert and Mohr (1967) reported that liquid medium with 1% coconut water caused death of Cattleya explants during the first three weeks of culture. When plantlets were grown on medium containing coconut water and sugar, optimal growth was obtained (Fig. 16C). The stimulating effect of coconut water was accounted for by Radley (1958) to the supply of diverse metabolites found in coconut water which were required for growth. Addition of 27. sucrose in the medium was found to be inhibitory to further multiplication of plbs. But sucrose was required for proliferation of the original explant tissue and also for root formation at a later stage. Lindemann (1970) showed that addition of

68 62 2% sucrose in medium caused Cattleya to form roots within ten days. The ill effect of sucrose was not established. It is suggested that possibly the presence of sucrose in medium for multiplication of plbs of Sarcanthine orchids may be due to osmotic effects of the external medium or connected with chlorophyll degradation as evidenced by the chlorosis of the tissues. For success and optimal growth of cultures, tissues had to be supplied with different media at various stages of development. The following media were found most suitable: (1) For proliferation of original explant tissue to callus, use liquid modified Vacin and Went. (2) For multiplication of plbs, both solid or liquid modified Vacin and Went without sucrose media may be used. (3) For maintaining plbs, use solid Vacin and Went without sucrose, (4) For plbs differentiation into plantlets, use solid Vacin and Went, (5) For good growth of plantlets, modified solid Vacin and Went is recommended. This observation demonstrated the concept that nutrient and growth substance requirements of a given tissue vary depending on the physiological state, as evidenced by the morphological-anatomical differentiation of the explant. For proliferation, coconut water was required. Though the exact constituent of coconut water was not determined, many workers (van Overbeek, 1941; Caplin and Steward, 1948) have reported that it contained various growth promoters.

69 63 principally kinetins (van Overbeek, 1941) which were known to cause cell division. The differentiation of roots from plbs required the addition of sucrose. In this way, growth and development of tissues could be manipulated by altering the components of the growth media.

70 SUMMARY The miniature monopodial orchids (Ascofinetia Twinkle, AscofInetia Cherry Blossum, Vandofinetia Premier, Neostylis Lou Sneary, Vascostylis Blue Fairy) were successfully mericloned on modified Vacin and Went medium. Explants from the young inflorescence were the most suitable material for culture, though apical buds and axillary buds near the apex could also be used. In the terminal and axillary bud culture, plbs originated from the cut surface of the scale-like leaves at the nodal area or from tissue of the young leaves. In inflorescence culture, plbs were derived from the epidermal cells. was initially slow but increased rapidly with time. Growth of plbs Three explant development patterns were observed; explant grew directly into plantlets, produced both shoots and plbs, or produced plbs only. Anatomical study revealed plbs consisted of undifferentiated mass of parenchymatous cells with areas of meristematic activity from which apical meristem and leaves originated. Vascular tissue was observed after leaves differentiated on the plbs. Addition of coconut water resulted in proliferation of tissues, while addition of sucrose resulted in necrosis of the undifferentiated plbs. Sucrose was required for root formation. For differentiation of plbs into plantlets and optimal growth of plantlets, solid Vacin and Went and solid modified Vacin and Went were found to be most suitable. This study demonstrated that monopodial miniature orchids did respond to meristem culture. Propagation of this group of orchids is not difficult, and has a similar commercial, potential as in Cymbidium and Dendrobium.
:382-383, Bertsch, W, 1967, A new frontier: Orchid propagation by meristem tissue culture.")

71 LITERATURE CITED Arditti, J, 1965, How good are tomato juice media for orchid seedling culture. Orchid Dig, 29(9): , Bertsch, W, 1967, A new frontier: Orchid propagation by meristem tissue culture. Am, Orchid Soc, Bull, 36:32-37, Bonnet, H, T, Jr, and J, G, Torrey, 1965, Chemical control of organ formation in root segments of Convovulus cultured in vitro. Plant Physiol, 40: , Caplin, S, M, and F, C, Steward, 1948, Effect of coconut milk on the growth of explants from carrot root. Science, 108: , Churchill, M, E,, E, A, Ball, and J, Arditti, 1970, Production of orchid plants from seedling leaf tips. Orchid Dig, 34: ,,,and,1971, Clonal propagation of orchids from leaf tips. Am, Orchid Soc, Bull, 40(2): , Ernest, R, 1967, Effect of select organic nutrient additives on growth in vitro of Phalaenopsis seedlings. Am, Orchid Soc, Bull, 36: ,, J, Arditti, and P, L, Healey, 1971, Carbohydrate physiology of orchid seedlings, II. Hydrolysis and effects oligosaccharides. Am, J, Botany, 58(9): , Ganapathy, P, S, 1969, Floral morphogenesis and flowering in aseptic cultures of Browallia demissa L, Biologia PI, 11: , Gautheret, R, J, 1955, The nutrition of plant tissue cultures, Ann, Rev, Plant Physiol, 6: , Halperin, W, 1966a, Alternative morphogenetic events in cell suspensions. Am, J, Botany, 53: , 1966b, Single cells, coconut milk and embryogenesis.in vitro. Science, 153: , Hayes, A, B, 1969, Mycorrhizal fungi and plant growth hormones. Am, Orchid Soc, Bull, 38: , Kent, A, E, and F, C, Steward, 1965, Morphogenesis in free cell cultures of carrot as affected by sequential treatments with Napthaleneacetic acid and with coconut milk. Am, J, Botany, 52:619p,
1-25. Konar, R. N. and A. Konar. 1966. Plantlet and flower formation in callus culture from Phlox drummandii Hook. Phytomorphology. 16:379-382. and K. Nataraja. 1969.")
72 66 Kim, K, K,, J, T, Kunisaki, and Y, Sagawa Shoot-tip culture of Dendrobiums. Am. Orchid Soc. Bull. 39(12); Knudson, L Non-symbiotic germination of orchid seeds. Bot. Gaz. 73(1)1-25. Konar, R. N. and A. Konar Plantlet and flower formation in callus culture from Phlox drummandii Hook. Phytomorphology. 16: and K. Nataraja Morphogenesis of isolated buds of Ranunculus scleratus L. in vitro. Acta Bot. Neerl. 18(6): Kotomori, S. and T. Murashige Some aspects of aseptic propagation of orchids. Am. Orchid Soc. Bull. 34: Lindemann, G. E. P Growth requirements for meristem culture of Gattleya. Hort. abstr. 38(4):8257., J, E. Gunckel, and 0. W. Davidson Meristem culture of Cattleya. Am. Orchid Soc. Bull. 39(11): Maheswari, S. C. and G. R. P. Gupta Production of adventitious embryoids in vitro from stem callus of Foeniculum vulgare. Planta. 67: Mohan Ram, H. Y. and M. Wadhi Morphogenic potentialities of flower buds of Kalanchoe pinnata Pers. in vitro. Ann. Bot. 32: Morel, G. M Producing virus-free Cymbidium. Am. Orchid Soc. Bull. 29: Tissue culture: a new means of clonal propagation of orchids. Am. Orchid Soc. Bull. 33: Clonal propagation of orchids by meristem culture. Cymbidium Soc, News, 20:3-11,. 1971, The principles of clonal propagation of orchids. Proceedings of the Sixth World Orchid Conference , Murashige, T, 1961, Suppression of shoot formation in cultured tobacco cells by gibberellic acid. Science. 134(3474):280., 1965, Effect of stem-elongation retardants and gibberellin on callus growth and organ formation in tobacco tissue culture. Physiol, Plantarum, 18: ,

73 Radley, M The distribution of substances similar to giberellic acid in higher plants. Ann. Bot. N. S. 22: Reinert, J., D. Backs, and M. Krosing Factors affecting embryogenesis in tissue cultures from domestic forms of Umbelifera. Planta. 68: Reinert, R. A. and H. C. Mohr Propagation of Cattleya by tissue cultures of lateral bud meristems. Proc. Am. Soc. Hort. Sci. 91: Sagawa, Y., T. Shoji, and T. Shoji Clonal propagation of Cymbidium through shoot meristem culture. Am. Orchid Soc. Bull. 35: _, and Clonal propagation of Dendrobiums through shoot meristem culture. Am. Orchid Soc. Bull. 36: and J. T. Kunisaki The possible use of tissue culture in horticulture. Proc. Am. Soc. Hort. Sci. 12: Scully, R. M. Jr Aspects of meristem culture in the Cattleya alliance. Am. Orchid Soc. Bull. 36: Sehgal, C. B Response of young inflorescences of Anethum graveolens L. to growth substances. Curr. Sci. 37(15): Shapiro, S Warm saffranin for plant tissues. Science Stabe, E. J., P. Laursen, and S. Buchner Medicinal plant tissue cultures. Proceedings International Conference on Plant Tissue Culture Steward, F. C., S. M. Caplin, and F. K. Millar Investigations on growth and metabolism of plant cells. I. New techniques for the investigation of metabolism, nutrition and growth in undifferentiated cells. Ann. Bot. 16:57-76., M. 0. Mapes, A. E. Kent, and R. D. Holsten Growth and development of cultured plant cells. Science. 143: Ueda, H. and H. Torikata Organogenesis in meristem cultures of cymbidiums. I. Studies on the effects of growth substances added to culture media under continuous illumination. J. Jap. Soc. Hort. Sci. 37:
74 68 Vacherot, M. 1966, Meristem tissue culture propagation of orchids. Proceedings of the Fifth World Orchid Conference, Vacin, E. and F, Went, 1949, Culture solution for orchid seedlings, Bot. Gaz. 110: van Overbeek, J,, M. E, Conklin, and A, F, Blakeslee, 1941, Factors in coconut milk essential for growth and development of young Datura embryos. Science, 94: , Wimber, D, E, 1963, Clonal multiplication of cymbidiums through tissue culture of the shoot meristem. Am, Orchid Soc, Bull, 32: ,, 1965, Additional observations on clonal multiplication of cymbidiums through culture of shoot meristem, Cymbidium Soc, News, 20(4):7-10, Yates, R, C, and J, T. Curtis, 1949c The effect of sucrose and other factors on the shoot-root ratio of orchid seedlings. Am, J, Botany, 36(5): ,
6 Production of Quality Planting Materials
 6 Production of Quality Planting Materials The lack of quality planting materials has become a major factor for limiting commercial cultivation of various orchids. There are six main techniques used for
6 Production of Quality Planting Materials The lack of quality planting materials has become a major factor for limiting commercial cultivation of various orchids. There are six main techniques used for
Phalaenopsis. Propagation Internodal. Sections
 J. Japan. Soc. Hort. Sci. 54 (3) : 379-387. 1985. New Means of Phalaenopsis Propagation with Internodal Sections of Flower Stalk' Yoshiyuki HOMMA' Faculty of Agriculture, and Kyoto Tadashi ASAHIRA University,
J. Japan. Soc. Hort. Sci. 54 (3) : 379-387. 1985. New Means of Phalaenopsis Propagation with Internodal Sections of Flower Stalk' Yoshiyuki HOMMA' Faculty of Agriculture, and Kyoto Tadashi ASAHIRA University,
Plant Regeneration via Organogenesis and Somatic Embryogenesis
 18 Plant Biotechnology: Practical Manual 2 Plant Regeneration via Organogenesis and Somatic Embryogenesis 2.1 BACKGROUND AND BASICS One can achieve plant regeneration in a test tube by using suitable culture
18 Plant Biotechnology: Practical Manual 2 Plant Regeneration via Organogenesis and Somatic Embryogenesis 2.1 BACKGROUND AND BASICS One can achieve plant regeneration in a test tube by using suitable culture
HIGHLY-EFFICIENT SOMATIC EMBRYOGENESIS FROM CELL SUSPENSION CULTURES OF PHALAENOPSIS ORCHIDS BY ADJUSTING CARBOHYDRATE SOURCES
 In Vitro Cell. Dev. Biol. Plant 39:635 639, November December 2003 q 2003 Society for In Vitro Biology 1054-5476/03 $18.00+0.00 DOI: 10.1079/IVP2003466 HIGHLY-EFFICIENT SOMATIC EMBRYOGENESIS FROM CELL
In Vitro Cell. Dev. Biol. Plant 39:635 639, November December 2003 q 2003 Society for In Vitro Biology 1054-5476/03 $18.00+0.00 DOI: 10.1079/IVP2003466 HIGHLY-EFFICIENT SOMATIC EMBRYOGENESIS FROM CELL
Callus induction and somatic embryogenesis of Phalaenopsis
 Plant Cell Reports (1998) 17: 446 450 Springer-Verlag 1998 Y. Ishii T. Takamura M. Goi M. Tanaka Callus induction and somatic embryogenesis of Phalaenopsis Received: 11 June 1997 / Revision received: 6
Plant Cell Reports (1998) 17: 446 450 Springer-Verlag 1998 Y. Ishii T. Takamura M. Goi M. Tanaka Callus induction and somatic embryogenesis of Phalaenopsis Received: 11 June 1997 / Revision received: 6
H. E. Sommer, H. Y. Wetzstein and N. Lee
 TISSUE CULTURE OF SWEETGUM (LIQUIDAMBAR STYRACIFLUA L.) H. E. Sommer, H. Y. Wetzstein and N. Lee Abstract.--An improved method for the tissue culture propagation of sweetgum (Liquidambar styraciflua L.)
TISSUE CULTURE OF SWEETGUM (LIQUIDAMBAR STYRACIFLUA L.) H. E. Sommer, H. Y. Wetzstein and N. Lee Abstract.--An improved method for the tissue culture propagation of sweetgum (Liquidambar styraciflua L.)
CHAPTER 2. IN VITRO REGENERATION OF Gerbera jamesonii Bolus Ex. Hook f. Previous research has proven that Gerbera jamesonii could successfully be
 CHAPTER 2 IN VITRO REGENERATION OF Gerbera jamesonii Bolus Ex. Hook f. 2.1 EXPERIMENTAL AIMS Previous research has proven that Gerbera jamesonii could successfully be propagated in vitro. Different types
CHAPTER 2 IN VITRO REGENERATION OF Gerbera jamesonii Bolus Ex. Hook f. 2.1 EXPERIMENTAL AIMS Previous research has proven that Gerbera jamesonii could successfully be propagated in vitro. Different types
Shoot Proliferation of Dendrobium Orchid with BAP and NAA
 Journal of Biological Sciences 3 (): 058-062, 2003 ISSN 727-3048 2003 Asian Network for Scientific Information Shoot Proliferation of Dendrobium Orchid with BAP and NAA 2 S.K. Talukder, K.M. Nasiruddin,
Journal of Biological Sciences 3 (): 058-062, 2003 ISSN 727-3048 2003 Asian Network for Scientific Information Shoot Proliferation of Dendrobium Orchid with BAP and NAA 2 S.K. Talukder, K.M. Nasiruddin,
DEPARTMENT OF BOTANY Guru Ghasidas Vishwavidyalaya, Bilaspur B. Sc. V Semester LBC 503 (Plant Tissue Culture)
") DEPARTMENT OF BOTANY Guru Ghasidas Vishwavidyalaya, Bilaspur B. Sc. V Semester LBC 503 (Plant Tissue Culture) Section A Multiple choice questions / Define the following terms 1 10 = 10 1. (d) Agrobacterium
DEPARTMENT OF BOTANY Guru Ghasidas Vishwavidyalaya, Bilaspur B. Sc. V Semester LBC 503 (Plant Tissue Culture) Section A Multiple choice questions / Define the following terms 1 10 = 10 1. (d) Agrobacterium
In Vitro Formation of Gerbera (Gerbera hybrida Hort.) Plantlets through Excised Scape Cultures
 Plantlets through Excised Scape Cultures") J. Japan. Soc. Hort. Sci. 52(1) : 45-50. 1983. In Vitro Formation of Gerbera (Gerbera hybrida Hort.) Plantlets through Excised Scape Cultures Chien-young CHU and Min-chang HUANG Department of Horticulture,
J. Japan. Soc. Hort. Sci. 52(1) : 45-50. 1983. In Vitro Formation of Gerbera (Gerbera hybrida Hort.) Plantlets through Excised Scape Cultures Chien-young CHU and Min-chang HUANG Department of Horticulture,
SOMATIC EMBRYOGENESIS OF DREPANOSTACHYUM FALCATUM AN IMPORTANT HILL BAMBOO-A RAPID MEANS OF MICROPROPAGATION
 SOMATIC EMBRYOGENESIS OF DREPANOSTACHYUM FALCATUM AN IMPORTANT HILL BAMBOO-A RAPID MEANS OF MICROPROPAGATION I.D.Arya, R. Sharma & Sarita Arya Forest Genetics & Tree Propagation Division, Arid Forest Research
SOMATIC EMBRYOGENESIS OF DREPANOSTACHYUM FALCATUM AN IMPORTANT HILL BAMBOO-A RAPID MEANS OF MICROPROPAGATION I.D.Arya, R. Sharma & Sarita Arya Forest Genetics & Tree Propagation Division, Arid Forest Research
RAPID PROPAGATION OF PHALAENOPSIS FROM FLORAL STALK-DERIVED LEAVES
 In Vitro Cell. Dev. Biol. Plant 38:168 172, March April 2002 q 2002 Society for In Vitro Biology 1054-5476/02 $10.00+0.00 DOI: 10.1079/IVP2001274 RAPID PROPAGATION OF PHALAENOPSIS FROM FLORAL STALK-DERIVED
In Vitro Cell. Dev. Biol. Plant 38:168 172, March April 2002 q 2002 Society for In Vitro Biology 1054-5476/02 $10.00+0.00 DOI: 10.1079/IVP2001274 RAPID PROPAGATION OF PHALAENOPSIS FROM FLORAL STALK-DERIVED
Effects of Complex Organic Extracts on Callus Growth and PLB Regeneration through Embryogenesis in the Doritaenopsis Orchid
 JARQ 37 (4), 229 235 (2003) http://www.jircas.affrc.go.jp Organic Extracts and Callus Growth and PLB Regenen of Doritaenopsis Effects of Complex Organic Extracts on Callus Growth and PLB Regenen through
JARQ 37 (4), 229 235 (2003) http://www.jircas.affrc.go.jp Organic Extracts and Callus Growth and PLB Regenen of Doritaenopsis Effects of Complex Organic Extracts on Callus Growth and PLB Regenen through
MICROPROPAGATION OF CHRYSANTHEMUM (CHRYSANTHEMUM MORIFOLIUM) USING SHOOT TIP AS EXPLANT
 USING SHOOT TIP AS EXPLANT") MICROPROPAGATION OF CHRYSANTHEMUM (CHRYSANTHEMUM MORIFOLIUM) USING SHOOT TIP AS EXPLANT R. Nalini Department of Biotechnology, Adhiparasakthi Agricultural College (Affiliated to TNAU) G. B. Nagar, Kalavai,
MICROPROPAGATION OF CHRYSANTHEMUM (CHRYSANTHEMUM MORIFOLIUM) USING SHOOT TIP AS EXPLANT R. Nalini Department of Biotechnology, Adhiparasakthi Agricultural College (Affiliated to TNAU) G. B. Nagar, Kalavai,
Plantlet Regeneration via Somatic Embryogenesis in Four Species of Crocus
 Plantlet Regeneration via Somatic Embryogenesis in Four Species of Crocus Roya Karamian Department of Biology, Faculty of Science Bu-Ali Sina University Hamadan Iran Keywords: meristems, micropropagation,
Plantlet Regeneration via Somatic Embryogenesis in Four Species of Crocus Roya Karamian Department of Biology, Faculty of Science Bu-Ali Sina University Hamadan Iran Keywords: meristems, micropropagation,
Plant regeneration through direct shoot bud formation from leaf cultures of Paphiopedilum orchids
 Plant Cell, Tissue and Organ Culture 76: 11 15, 2004. 2003 Kluwer Academic Publishers. Printed in the Netherlands. 11 Plant regeneration through direct shoot bud formation from leaf cultures of Paphiopedilum
Plant Cell, Tissue and Organ Culture 76: 11 15, 2004. 2003 Kluwer Academic Publishers. Printed in the Netherlands. 11 Plant regeneration through direct shoot bud formation from leaf cultures of Paphiopedilum
AVOCADO CALLUS AND BUD CULTURE
 Proc. Fla. State Hort. Soc. 96:181-182. 1983. AVOCADO CALLUS AND BUD CULTURE M. J. Young University of Florida, IF AS, Fruit Crops Department, Gainesville, FL 32611 Additional index words, tissue culture,
Proc. Fla. State Hort. Soc. 96:181-182. 1983. AVOCADO CALLUS AND BUD CULTURE M. J. Young University of Florida, IF AS, Fruit Crops Department, Gainesville, FL 32611 Additional index words, tissue culture,
Utilization of Embryogenic Cell Cultures for the Mass Production of Bulblets in Lilies
 Utilization of Embryogenic Cell Cultures for the Mass Production of Bulblets in Lilies Sun Ki Kim and Byung Joon Ahn College of Bioresources Science Dankook University Cheonan 330-714 Korea Keywords: Oriental
Utilization of Embryogenic Cell Cultures for the Mass Production of Bulblets in Lilies Sun Ki Kim and Byung Joon Ahn College of Bioresources Science Dankook University Cheonan 330-714 Korea Keywords: Oriental
THE ROLE OF TISSUE CULTURE IN THE AVOCADO PLANT IMPROVEMENT SCHEME
 South African Avocado Growers Association Yearbook 1984. 7:25-26 THE ROLE OF TISSUE CULTURE IN THE AVOCADO PLANT IMPROVEMENT SCHEME Review DOROTHEA D NEL AND JM KOTZÉ DEPT OF MICROBIOLOGY AND PLANT PATHOLOGY,
South African Avocado Growers Association Yearbook 1984. 7:25-26 THE ROLE OF TISSUE CULTURE IN THE AVOCADO PLANT IMPROVEMENT SCHEME Review DOROTHEA D NEL AND JM KOTZÉ DEPT OF MICROBIOLOGY AND PLANT PATHOLOGY,
Micropropagation Scheme of Curcuma alismatifolia Gagnep.
 Micropropagation Scheme of Curcuma alismatifolia Gagnep. N. Toppoonyanont, S. Chongsang, S. Chujan, S. Somsueb and P. Nuamjaroen Department of Biology, Faculty of Science, Maejo University Chiang Mai 50290
Micropropagation Scheme of Curcuma alismatifolia Gagnep. N. Toppoonyanont, S. Chongsang, S. Chujan, S. Somsueb and P. Nuamjaroen Department of Biology, Faculty of Science, Maejo University Chiang Mai 50290
Chapter 1 Introduction
 Chapter 1 Introduction Experimental systems based on plant cell and tissue culture are characterized by the use of isolated parts of, called ex, obtained from an intact plant body and kept on, or in a
Chapter 1 Introduction Experimental systems based on plant cell and tissue culture are characterized by the use of isolated parts of, called ex, obtained from an intact plant body and kept on, or in a
Micropropagation of a terrestrial Cymbidium species using rhizomes developed from seeds and pseudobulbs
 Plant Cell, Tissue and Organ Culture 22: 237-244, 1990. 1990 Kluwer Academic Publishers. Printed in the Netherlands. Micropropagation of a terrestrial Cymbidium species using rhizomes developed from seeds
Plant Cell, Tissue and Organ Culture 22: 237-244, 1990. 1990 Kluwer Academic Publishers. Printed in the Netherlands. Micropropagation of a terrestrial Cymbidium species using rhizomes developed from seeds
EFFECT OF BENZYLAMINO PURINE AND NAPHTHALENE ACETIC ACID ON CALLUS AND PROTOCORM FORMATION OF DENDROBIUM CV. BANYAT PINK
 Journal of Cell and Tissue Research Vol. 13(3) 3977-3981 (2013) (Available online at www.tcrjournals.com) ISSN: 0973-0028; E-ISSN: 0974-0910 Original Article EFFECT OF BENZYLAMINO PURINE AND NAPHTHALENE
Journal of Cell and Tissue Research Vol. 13(3) 3977-3981 (2013) (Available online at www.tcrjournals.com) ISSN: 0973-0028; E-ISSN: 0974-0910 Original Article EFFECT OF BENZYLAMINO PURINE AND NAPHTHALENE
Gregor Mendel Foundation Proceedings 2007:
 Gregor Mendel Foundation Proceedings 2007: 54-61. Effect of the explants from different aged mother plant on callus induction and direct regeneration in jatropha(jatropha curcas L.) Pranesh K. J 1, Gururaja
Gregor Mendel Foundation Proceedings 2007: 54-61. Effect of the explants from different aged mother plant on callus induction and direct regeneration in jatropha(jatropha curcas L.) Pranesh K. J 1, Gururaja
In vitro Conservation of Rose Coloured Leadwort: Effect of Mannitol on Growth of Plantlets
 Kasetsart J. (Nat. Sci.) 38 : 97-12 (24) In vitro Conservation of Rose Coloured Leadwort: Effect of Mannitol on Growth of Plantlets Rommanee Charoensub and Salak Phansiri ABSTRACT In vitro study of manitol
Kasetsart J. (Nat. Sci.) 38 : 97-12 (24) In vitro Conservation of Rose Coloured Leadwort: Effect of Mannitol on Growth of Plantlets Rommanee Charoensub and Salak Phansiri ABSTRACT In vitro study of manitol
Original Papers Plant Tissue Culture Letters, 11(3), (1994) INouE**, Koji NoMuRA***, Seiko TAKAHASHi**, OsHIMA* and Kiyoshi MASUDA**
, (1994) INouE**, Koji NoMuRA***, Seiko TAKAHASHi**, OsHIMA* and Kiyoshi MASUDA**") Original Papers Plant Tissue Culture Letters, 11(3), 184-190 (1994) An Efficient Procedure for Plant Regeneration from Long-term-cultured Callus Tissue of Hybrid Poplar (Populus sieboldii Miq. X Populus
Original Papers Plant Tissue Culture Letters, 11(3), 184-190 (1994) An Efficient Procedure for Plant Regeneration from Long-term-cultured Callus Tissue of Hybrid Poplar (Populus sieboldii Miq. X Populus
Keywords: Agarwood, satalum, resin, micropropagation, tok
 Application of Tissue Culture Techniques in Woody Species Conservation, Improvement and Development in Vietnam: Agarwood (Aquilaria crassna Pierre ex LeComte) via tip Culture Tran Van Minh Institute of
Application of Tissue Culture Techniques in Woody Species Conservation, Improvement and Development in Vietnam: Agarwood (Aquilaria crassna Pierre ex LeComte) via tip Culture Tran Van Minh Institute of
In Vitro Microcorm Formation in Saffron (Crocus sativus L.)
") In Vitro Microcorm Formation in Saffron (Crocus sativus L.) W. Raja, G. Zaffer, S.A. Wani Division of Plant Breeding & Genetics Sher-e-Kashmir University of Agricultural Sciences and Technology of Kashmir
In Vitro Microcorm Formation in Saffron (Crocus sativus L.) W. Raja, G. Zaffer, S.A. Wani Division of Plant Breeding & Genetics Sher-e-Kashmir University of Agricultural Sciences and Technology of Kashmir
Title: Development of Micropropagation and Acclimation Protocols for the Commercialization of a New Bonsai Ornamaental Tree for the California Market.
 Title: Development of Micropropagation and Acclimation Protocols for the Commercialization of a New Bonsai Ornamaental Tree for the California Market. Authors: Dan E. Parfitt 1, Helen M. Chan 2, and Ali
Title: Development of Micropropagation and Acclimation Protocols for the Commercialization of a New Bonsai Ornamaental Tree for the California Market. Authors: Dan E. Parfitt 1, Helen M. Chan 2, and Ali
IN VITRO GROWTH AND DEVELOPMENT OF DENDROBIUM HYBRID ORCHID. H. KHATUN 1, M. M. KHATUN 2, M. S. BISWAS 3 M. R. KABIR 4 AND M. AL-AMIN 5 Abstract
 ISSN 0258-7122 Bangladesh J. Agril. Res. 35(3) : 507-514, September 2010 IN VITRO GROWTH AND DEVELOPMENT OF DENDROBIUM HYBRID ORCHID H. KHATUN 1, M. M. KHATUN 2, M. S. BISWAS 3 M. R. KABIR 4 AND M. AL-AMIN
ISSN 0258-7122 Bangladesh J. Agril. Res. 35(3) : 507-514, September 2010 IN VITRO GROWTH AND DEVELOPMENT OF DENDROBIUM HYBRID ORCHID H. KHATUN 1, M. M. KHATUN 2, M. S. BISWAS 3 M. R. KABIR 4 AND M. AL-AMIN
MULTIPLE SHOOT REGENERATION IN DENDROBIUM FIMBRIATUM HOOK AN ORNAMENTAL ORCHID ABSTRACT
 Kabir et al., The Journal of Animal & Plant Sciences, 23(4): 13, Page: J. 1140-1145 Anim. Plant Sci. 23(4):13 ISSN: 1018-7081 MULTIPLE SHOOT REGENERATION IN DENDROBIUM FIMBRIATUM HOOK AN ORNAMENTAL ORCHID
Kabir et al., The Journal of Animal & Plant Sciences, 23(4): 13, Page: J. 1140-1145 Anim. Plant Sci. 23(4):13 ISSN: 1018-7081 MULTIPLE SHOOT REGENERATION IN DENDROBIUM FIMBRIATUM HOOK AN ORNAMENTAL ORCHID
VETIVER PROPAGATION. Nurseries and Large Scale Propagation. Dr Paul Truong Veticon Consulting Brisbane, Australia
 VETIVER PROPAGATION Nurseries and Large Scale Propagation Dr Paul Truong Veticon Consulting Brisbane, Australia 1. INTRODUCTION The Vetiver Network promotes the use of sterile vetiver cultivar to avoid
VETIVER PROPAGATION Nurseries and Large Scale Propagation Dr Paul Truong Veticon Consulting Brisbane, Australia 1. INTRODUCTION The Vetiver Network promotes the use of sterile vetiver cultivar to avoid
Plant Propagation-The Union of
 Plant Propagation-The Union of Art and Science I would like to start my presentation with a word of sincere appreciation to arboreta and botantical gardens in general and the Arnold Arboretum in particular
Plant Propagation-The Union of Art and Science I would like to start my presentation with a word of sincere appreciation to arboreta and botantical gardens in general and the Arnold Arboretum in particular
In vitro propagation of Phaleonopsis hybrid Little gem by culturing apical part and axillary bud of flower stalk
 J Plant Biotechnol (2016) 43:438 443 DOI:https://doi.org/10.5010/JPB.2016.43.4.438 ISSN 1229-2818 (Print) ISSN 2384-1397 (Online) Research Article In vitro propagation of Phaleonopsis hybrid Little gem
J Plant Biotechnol (2016) 43:438 443 DOI:https://doi.org/10.5010/JPB.2016.43.4.438 ISSN 1229-2818 (Print) ISSN 2384-1397 (Online) Research Article In vitro propagation of Phaleonopsis hybrid Little gem
Literature on micropropagation of awa (Piper
 Biotechnology Jan. 2003 BI-4 Micropropagation of Awa (Kava, Piper methysticum) J. Kunisaki, A. Araki, and Y. Sagawa Department of Tropical Plant and Soil Sciences Literature on micropropagation of awa
Biotechnology Jan. 2003 BI-4 Micropropagation of Awa (Kava, Piper methysticum) J. Kunisaki, A. Araki, and Y. Sagawa Department of Tropical Plant and Soil Sciences Literature on micropropagation of awa
Effect of Cytokinins on Multiple Shoot Regeneration from Leaf Derived Callus of Inula
 Plant Tissue Cult. & Biotech. 27(2): 189 194, 2017 (December) PTC&B Effect of Cytokinins on Multiple Shoot Regeneration from Leaf Derived Callus of Inula royleana DC. Samar Amin*, Zahoor A Kaloo and Seema
Plant Tissue Cult. & Biotech. 27(2): 189 194, 2017 (December) PTC&B Effect of Cytokinins on Multiple Shoot Regeneration from Leaf Derived Callus of Inula royleana DC. Samar Amin*, Zahoor A Kaloo and Seema
Asexual Reproduction
 Asexual Reproduction Methods of Vegetative Propagation Cutting Grafting and budding Layering Specialised stems and roots Cutting Propagation by Cutting e.g. spruce, pine, juniper, citrus, olive, apple
Asexual Reproduction Methods of Vegetative Propagation Cutting Grafting and budding Layering Specialised stems and roots Cutting Propagation by Cutting e.g. spruce, pine, juniper, citrus, olive, apple
Summary and conclusion
 Summary and conclusion 6.1 Chemical sterilization of Explants Surface sterilization of explants collected from mature trees is necessary before initiation of any in vitro culture. In the present study
Summary and conclusion 6.1 Chemical sterilization of Explants Surface sterilization of explants collected from mature trees is necessary before initiation of any in vitro culture. In the present study
Micro propagation of sugarcane (Saccharum officinarum L.) through auxiliary buds
 through auxiliary buds") Micro propagation of sugarcane (Saccharum officinarum L.) through auxiliary buds P S \Varakagoda, S Subasinghe, D L C Kumari and T S Neththikumara Department of Crop Science, Faculty of Agriculture, University
Micro propagation of sugarcane (Saccharum officinarum L.) through auxiliary buds P S \Varakagoda, S Subasinghe, D L C Kumari and T S Neththikumara Department of Crop Science, Faculty of Agriculture, University
Plant Propagation PLS 3223/5222
 Plant Propagation PLS 3223/5222 Guest Web Lecture Dr. Michael Kane Environmental Horticulture Department PLANT MICROPROPAGATION 1 Micropropagation Rapid clonal in vitro ( in glass ) propagation of plants
Plant Propagation PLS 3223/5222 Guest Web Lecture Dr. Michael Kane Environmental Horticulture Department PLANT MICROPROPAGATION 1 Micropropagation Rapid clonal in vitro ( in glass ) propagation of plants
Effect of BA and 2iP on Shoot Proliferation and Somaclonal Variation of Gardenia jasminoides Ellis in vitro Culture
 R ESEARCH ARTICLE ScienceAsia 27 (2001) : 137-141 Effect of BA and 2iP on Shoot Proliferation and Somaclonal Variation of Gardenia jasminoides Ellis in vitro Culture Ngarmnij Chuenboonngarm a,b, Suvimon
R ESEARCH ARTICLE ScienceAsia 27 (2001) : 137-141 Effect of BA and 2iP on Shoot Proliferation and Somaclonal Variation of Gardenia jasminoides Ellis in vitro Culture Ngarmnij Chuenboonngarm a,b, Suvimon
Grafting Morphology and Physiology Text Pages:
 65 Grafting Morphology and Physiology Text Pages: 415 432. Objectives: 1. Be able to describe and explain the terms used in grafting. 2. Be able to describe and summarize the reasons for grafting plants.
65 Grafting Morphology and Physiology Text Pages: 415 432. Objectives: 1. Be able to describe and explain the terms used in grafting. 2. Be able to describe and summarize the reasons for grafting plants.
A micropropagation system for Eucalyptus dunnii Eucalyptus sp
 A micropropagation system for Eucalyptus dunnii Eucalyptus sp M. Fantini Jr., M.E. Cortezzi Graça To cite this version: M. Fantini Jr., M.E. Cortezzi Graça. A micropropagation system for Eucalyptus dunnii
A micropropagation system for Eucalyptus dunnii Eucalyptus sp M. Fantini Jr., M.E. Cortezzi Graça To cite this version: M. Fantini Jr., M.E. Cortezzi Graça. A micropropagation system for Eucalyptus dunnii
Question Bank Vegetative Propagation
 Question Bank Vegetative Propagation 1. Define vegetative reproduction. Ans. Vegetative reproduction is the formation of a new individual from any vegetative part of the plant body. 2. Mention any two
Question Bank Vegetative Propagation 1. Define vegetative reproduction. Ans. Vegetative reproduction is the formation of a new individual from any vegetative part of the plant body. 2. Mention any two
Polysiphonia. Life cycle of Polysiphonia Dr. Purnima Saikia, Asst.Prof Dept of Botany2017
 Polysiphonia Life cycle of Polysiphonia Dr. Purnima Saikia, Asst.Prof Dept of Botany2017 Polysiphonia thallus Development of female gametophyte MALE SEX ORGANS Cystocarp with carposporangium Mature Cystocarp
Polysiphonia Life cycle of Polysiphonia Dr. Purnima Saikia, Asst.Prof Dept of Botany2017 Polysiphonia thallus Development of female gametophyte MALE SEX ORGANS Cystocarp with carposporangium Mature Cystocarp
Micropropagation of Sterile and Non-Flowering Nicotiana Lines
 2008 The Japan Mendel Society Cytologia 73(1): 9 13, 2008 Micropropagation of Sterile and Non-Flowering Nicotiana Lines Sarala K.*, Rao R. V. S., Murthy T. G. K. and Satyavani J. V. R. Central Tobacco
2008 The Japan Mendel Society Cytologia 73(1): 9 13, 2008 Micropropagation of Sterile and Non-Flowering Nicotiana Lines Sarala K.*, Rao R. V. S., Murthy T. G. K. and Satyavani J. V. R. Central Tobacco
Review on Adventitious Bud Formation in Lachenalia
 Review on Adventitious Bud Formation in Lachenalia J.G. Niederwieser and A.M. Ndou ARC-Roodeplaat Vegetable and Ornamental Plant Institute Private Bag X293 Pretoria, 0001 South Africa Tel: 27-12 841 9609
Review on Adventitious Bud Formation in Lachenalia J.G. Niederwieser and A.M. Ndou ARC-Roodeplaat Vegetable and Ornamental Plant Institute Private Bag X293 Pretoria, 0001 South Africa Tel: 27-12 841 9609
The application of leafy explant micropropagation protocol in enhancing the multiplication ef ciency of Alstroemeria
 Scientia Horticulturae 85 (2000) 307±318 The application of leafy explant micropropagation protocol in enhancing the multiplication ef ciency of Alstroemeria Hsueh-Shih Lin 1, Marjo J. De Jeu *, Evert
Scientia Horticulturae 85 (2000) 307±318 The application of leafy explant micropropagation protocol in enhancing the multiplication ef ciency of Alstroemeria Hsueh-Shih Lin 1, Marjo J. De Jeu *, Evert
In vitro Plant Regeneration of Withania somnifera. Ujjwala Supe, Fanisha Dhote and M.G. Roymon
 Plant Tissue Cult. & Biotech. 16(2): 111-115, 2006 (December) - Short communication PTC&B In vitro Plant Regeneration of Withania somnifera Ujjwala Supe, Fanisha Dhote and M.G. Roymon Plant Tissue Culture
Plant Tissue Cult. & Biotech. 16(2): 111-115, 2006 (December) - Short communication PTC&B In vitro Plant Regeneration of Withania somnifera Ujjwala Supe, Fanisha Dhote and M.G. Roymon Plant Tissue Culture
Plant Tissue Culture. Dr. Alain Lemansour UAE University Date Palm Development Research Unit Dept.
 Plant Tissue Culture By Dr. Alain Lemansour UAE University Date Palm Development Research Unit Dept. What is it? Tissue culture is the term used for the process of growing cells artificially in the laboratory
Plant Tissue Culture By Dr. Alain Lemansour UAE University Date Palm Development Research Unit Dept. What is it? Tissue culture is the term used for the process of growing cells artificially in the laboratory
INFLUENCE OF PLANT GROWTH REGULATORS ON IN VITRO CLONAL PROPAGATION OF DENDROBIUM SONIA EARSAKUL
 INFLUENCE OF PLANT GROWTH REGULATORS ON IN VITRO CLONAL PROPAGATION OF DENDROBIUM SONIA EARSAKUL Priya Kumari, I.* Sabina George, T. & Rajmohan, K 1 Department of Pomology and Floriculture, College of
INFLUENCE OF PLANT GROWTH REGULATORS ON IN VITRO CLONAL PROPAGATION OF DENDROBIUM SONIA EARSAKUL Priya Kumari, I.* Sabina George, T. & Rajmohan, K 1 Department of Pomology and Floriculture, College of
ORGANOGENESIS IN CHRYSANTHEMUM MORIFOLIUM RAMAT (CULTIVAR ROMICA ) CALLUS CULTURES SMARANDA VÂNTU
 CALLUS CULTURES SMARANDA VÂNTU") Analele ştiinţifice ale Universităţii Al. I. Cuza Iaşi Tomul LII, s. II a. Biologie vegetală, 006 ORGANOGENESIS IN CHRYSANTHEMUM MORIFOLIUM RAMAT (CULTIVAR ROMICA ) CALLUS CULTURES SMARANDA VÂNTU Abstract:
Analele ştiinţifice ale Universităţii Al. I. Cuza Iaşi Tomul LII, s. II a. Biologie vegetală, 006 ORGANOGENESIS IN CHRYSANTHEMUM MORIFOLIUM RAMAT (CULTIVAR ROMICA ) CALLUS CULTURES SMARANDA VÂNTU Abstract:
MICROPROPAGATION OF RICE (ORYZA SATIVA L. CV SWAT-II) THROUGH SOMATIC EMBRYOGENESIS
 THROUGH SOMATIC EMBRYOGENESIS") Pak. J. Bot., 37(2): 237-242, 2005. MICROPROPAGATION OF RICE (ORYZA SATIVA L. CV SWAT-II) THROUGH SOMATIC EMBRYOGENESIS IHSAN ILAHI, SHAZIA BANO, MUSARRAT JABEEN AND FAZAL RAHIM Department of Botany, University
Pak. J. Bot., 37(2): 237-242, 2005. MICROPROPAGATION OF RICE (ORYZA SATIVA L. CV SWAT-II) THROUGH SOMATIC EMBRYOGENESIS IHSAN ILAHI, SHAZIA BANO, MUSARRAT JABEEN AND FAZAL RAHIM Department of Botany, University
ACHIEVEMENT LEVEL DESCRIPTORS
 ACHIEVEMENT LEVEL DESCRIPTORS FOR THE PLANT SYSTEMS ASSESSMENT Please note: Students performing at the Meets Expectations level also meet all standards at the Approaches Expectations level, and students
ACHIEVEMENT LEVEL DESCRIPTORS FOR THE PLANT SYSTEMS ASSESSMENT Please note: Students performing at the Meets Expectations level also meet all standards at the Approaches Expectations level, and students
TISSUE CULTURE II. Organogenesis. PlSc 300 LAB Learn tissue culture techniques that promote organ formation.
 76 TISSUE CULTURE II Organogenesis PlSc 300 LAB 11 REFERENCE: Text: 663 666; 706 712; 717 718. OBJECTIVES: 1. Learn tissue culture techniques that promote organ formation. 2. Practice making leaf and shoot
76 TISSUE CULTURE II Organogenesis PlSc 300 LAB 11 REFERENCE: Text: 663 666; 706 712; 717 718. OBJECTIVES: 1. Learn tissue culture techniques that promote organ formation. 2. Practice making leaf and shoot
Studies on Vegetative Propagation ~.f Tulips Regeneration of Bulblets in Bulb Scale Segments Culturt. 1 in vitro
 J. Japan. Soc. Hort. Sci. 49(2) : 235-24u. ' QR0. Studies on Vegetative Propagation ~.f Tulips IV. Regeneration of Bulblets in Bulb Scale Segments Culturt. 1 in vitro Yoshlo NISHIUCHI Hokkaido University
J. Japan. Soc. Hort. Sci. 49(2) : 235-24u. ' QR0. Studies on Vegetative Propagation ~.f Tulips IV. Regeneration of Bulblets in Bulb Scale Segments Culturt. 1 in vitro Yoshlo NISHIUCHI Hokkaido University
RETRACTED. Key words: asymbiotic culture, flasking, horticultural orchid, in vitro propagation
 Micropropagation of Dendrobium chrysanthum Wall. 81 FOLIA HORTICULTURAE Ann. 18/1, 2006, 81-90 Green Pod Culture and Rapid Micropropagation of Dendrobium chrysanthum Wall. A Horticultural and Medicinal
Micropropagation of Dendrobium chrysanthum Wall. 81 FOLIA HORTICULTURAE Ann. 18/1, 2006, 81-90 Green Pod Culture and Rapid Micropropagation of Dendrobium chrysanthum Wall. A Horticultural and Medicinal
Increasing the growth rate by any means decreases the juvenile period
 FRUITING Growth stages Juvenile phase: The non-flowering period after seed germination. Morphological differences Leaf shape and thickness Ability to form adventitious roots Seedling can not be induced
FRUITING Growth stages Juvenile phase: The non-flowering period after seed germination. Morphological differences Leaf shape and thickness Ability to form adventitious roots Seedling can not be induced
Temjensangba and Chitta Ranjan Deb*
 Indian Journal of Biotechnology Vol 5, April 2006, pp 223-228 Effect of different factors on non-symbiotic seed germination, formation of protocorm-like bodies and plantlet morphology of Cleisostoma racemiferum
Indian Journal of Biotechnology Vol 5, April 2006, pp 223-228 Effect of different factors on non-symbiotic seed germination, formation of protocorm-like bodies and plantlet morphology of Cleisostoma racemiferum
Regeneration of plantlets from in vitro Root and Leaf culture of Vandaceous orchid, Ascocentrum ampullaceum (Roxb.) Schlter
 Schlter") International Journal of Agriculture, Environment and Biotechnology Citation: IJAEB: 9(4): 659-666 August 2016 DOI Number: 10.5958/2230-732X.2016.00086.3 2016 New Delhi Publishers. All rights reserved
International Journal of Agriculture, Environment and Biotechnology Citation: IJAEB: 9(4): 659-666 August 2016 DOI Number: 10.5958/2230-732X.2016.00086.3 2016 New Delhi Publishers. All rights reserved
IN VITRO BUD CULTURE OF KINNOW TREE
 Pak. J. Bot., 38(3): 597-601, 2006. IN VITRO BUD CULTURE OF KINNOW TREE Nuclear Institute for Agriculture Biology (NIAB), Faisalabad. Abstract Tissues from field grown trees have contamination problems
Pak. J. Bot., 38(3): 597-601, 2006. IN VITRO BUD CULTURE OF KINNOW TREE Nuclear Institute for Agriculture Biology (NIAB), Faisalabad. Abstract Tissues from field grown trees have contamination problems
REGENERATION OF PLANTLETS FROM IN VITRO LEAF CULTURE OF RENADES ARUNODAY HYBRID ( AERIDES ROSEA LODDIGES EX PAXT. X RENANTHERA IMSCHOOTIANA ROLFE)
") J. Orchid Soc. ndia, 13(1-2): 19-24, 1999 REGENERATON OF PLANTLETS FROM N VTRO LEAF CULTURE OF RENADES ARUNODAY HYBRD ( AERDES ROSEA LODDGES EX PAXT. X RENANTHERA MSCHOOTANA ROLFE) ( S.K. Sinha* and Sadanand
J. Orchid Soc. ndia, 13(1-2): 19-24, 1999 REGENERATON OF PLANTLETS FROM N VTRO LEAF CULTURE OF RENADES ARUNODAY HYBRD ( AERDES ROSEA LODDGES EX PAXT. X RENANTHERA MSCHOOTANA ROLFE) ( S.K. Sinha* and Sadanand
International Journal of Pharma and Bio Sciences DIRECT REGENERATION FROM EMBRYO CULTURES OF LYCOPERSICON ESCULENTUM MILL CV PUSA RUBY ABSTRACT
 Research Article Plant science International Journal of Pharma and Bio Sciences ISSN 0975-6299 DIRECT REGENERATION FROM EMBRYO CULTURES OF LYCOPERSICON ESCULENTUM MILL CV PUSA RUBY P. KARUNAKAR RAO Department
Research Article Plant science International Journal of Pharma and Bio Sciences ISSN 0975-6299 DIRECT REGENERATION FROM EMBRYO CULTURES OF LYCOPERSICON ESCULENTUM MILL CV PUSA RUBY P. KARUNAKAR RAO Department
Researches regarding bitter melon (Momordica charantia) in vitro regeneration
 in vitro regeneration") Volume 14(3), 75-8, 21 JOURNAL of Horticulture, Forestry and Biotechnology Researches regarding bitter melon (Momordica charantia) in vitro regeneration Franţ Alexandra 1*, Botău Dorica 1 1 Banat s University
Volume 14(3), 75-8, 21 JOURNAL of Horticulture, Forestry and Biotechnology Researches regarding bitter melon (Momordica charantia) in vitro regeneration Franţ Alexandra 1*, Botău Dorica 1 1 Banat s University
CMG GardenNotes #659 Understanding Tree Roots. Functions of Tree Roots. Support\anchorage
 CMG GardenNotes #659 Understanding Tree Roots Outline: Functions of Tree Roots, page 1 Describing Tree Roots, page 2 Conditions That Can Adversely Affect Roots, page 4 Other Common Root Issues, page 5
CMG GardenNotes #659 Understanding Tree Roots Outline: Functions of Tree Roots, page 1 Describing Tree Roots, page 2 Conditions That Can Adversely Affect Roots, page 4 Other Common Root Issues, page 5
Adult Plants and Juvenile Seedlings of Persimmon (Diospyros kaki L.)
") J. Japan. Soc. Hort. Sci. 63(3) : 537-541. 1994. Comparison of Growth Rooting Characteristics of Micropropagated Adult Plants Juvenile Seedlings of Persimmon (Diospyros kaki L.) Ryutaro Tao, Jun Ito Akira
J. Japan. Soc. Hort. Sci. 63(3) : 537-541. 1994. Comparison of Growth Rooting Characteristics of Micropropagated Adult Plants Juvenile Seedlings of Persimmon (Diospyros kaki L.) Ryutaro Tao, Jun Ito Akira
2. What are the advantages and disadvantages of plant tissue culture?
 UNIT VI - PLANT TECHNOLOGIES Lesson 2: Plant Tissue Culture Competency/Objective: Explain the process of tissue culture. Study Questions References 1. What is plant tissue culture? 2. What are the advantages
UNIT VI - PLANT TECHNOLOGIES Lesson 2: Plant Tissue Culture Competency/Objective: Explain the process of tissue culture. Study Questions References 1. What is plant tissue culture? 2. What are the advantages
IN VITRO SHOOT MULTIPLICATION AND CALLUS INDUCTION IN GLADIOLUS HYBRIDUS HORT.
 Pak. J. Bot., 40(2): 517-522, 2008. IN VITRO SHOOT MULTIPLICATION AND CALLUS INDUCTION IN GLADIOLUS HYBRIDUS HORT. FAHEEM AFTAB, MEMOONA ALAM AND HUMERA AFRASIAB Department of Botany, University of the
Pak. J. Bot., 40(2): 517-522, 2008. IN VITRO SHOOT MULTIPLICATION AND CALLUS INDUCTION IN GLADIOLUS HYBRIDUS HORT. FAHEEM AFTAB, MEMOONA ALAM AND HUMERA AFRASIAB Department of Botany, University of the
HOSTA MULTIPLICATION KIT
 HOSTA MULTIPLICATION KIT Product No. H411 PhytoTechnology Laboratories Mailing Address: P.O. Box 12205, Shawnee Mission, KS 66282-2205 Phone: 1-888-749-8682 (1-913-341-5343 Outside the USA & Canada) Fax:
HOSTA MULTIPLICATION KIT Product No. H411 PhytoTechnology Laboratories Mailing Address: P.O. Box 12205, Shawnee Mission, KS 66282-2205 Phone: 1-888-749-8682 (1-913-341-5343 Outside the USA & Canada) Fax:
The preferred carbohydrate in plant cell culture media is sucrose. Glucose and fructose may be substituted in some cases, glucose being as
 Introduction One of the most important factors governing the growth and morphogenesis of plant tissues in culture is the composition of the culture medium. The basic nutrient requirements of cultured plant
Introduction One of the most important factors governing the growth and morphogenesis of plant tissues in culture is the composition of the culture medium. The basic nutrient requirements of cultured plant
B. C. D. B. C. D. B. C. D. B. C. D.
 Student Name: Teacher: Date: Test: 9_12 Agriculture AP41 - Horticulture I Test 1 Description: Unit C-Plant Physiology District: Wake County Form: 501 1. Water loss from leaf surface through evaporation
Student Name: Teacher: Date: Test: 9_12 Agriculture AP41 - Horticulture I Test 1 Description: Unit C-Plant Physiology District: Wake County Form: 501 1. Water loss from leaf surface through evaporation
4. VEGETATIVE PROPAGATION
 The Division of Subtropical Agriculture. The Volcani Institute of Agricultural Research 1960-1969. Section B. Avocado. Pg 47-54. 4. VEGETATIVE PROPAGATION a. Rooting of Leaf-bearing Cuttings - A. Kadman
The Division of Subtropical Agriculture. The Volcani Institute of Agricultural Research 1960-1969. Section B. Avocado. Pg 47-54. 4. VEGETATIVE PROPAGATION a. Rooting of Leaf-bearing Cuttings - A. Kadman
Efficient micropropagation of Vanilla planifolia Andr. under influence of thidiazuron, zeatin and coconut milk
 Indian Journal of Biotechnology Vol 3, January 2004, pp. 113-118 Efficient micropropagation of Vanilla planifolia Andr. under influence of thidiazuron, zeatin and coconut milk P Giridhar and G A Ravishankar*
Indian Journal of Biotechnology Vol 3, January 2004, pp. 113-118 Efficient micropropagation of Vanilla planifolia Andr. under influence of thidiazuron, zeatin and coconut milk P Giridhar and G A Ravishankar*
Environmental and Genotypic Effects on the Growth Rate. of in Vitro Cassava Plantlet
 1 Environmental and Genotypic Effects on the Growth Rate of in Vitro Cassava Plantlet By Sunday E. Aladele National Centre for Genetic Resources and Biotechnology, Ibadan, Nigeria Abstract Two cassava
1 Environmental and Genotypic Effects on the Growth Rate of in Vitro Cassava Plantlet By Sunday E. Aladele National Centre for Genetic Resources and Biotechnology, Ibadan, Nigeria Abstract Two cassava
Clonal Propagation of White Flowered Hybrid Cattleya through in Vitro Culture of Thin Cell Layers from Young Shoots of Mature Plants
 Orchid Science and Biotechnology 27 Global Science Books Clonal Propagation of White Flowered Hybrid Cattleya through in Vitro Culture of Thin Cell Layers from Young Shoots of Mature Plants Pinaki Sinha
Orchid Science and Biotechnology 27 Global Science Books Clonal Propagation of White Flowered Hybrid Cattleya through in Vitro Culture of Thin Cell Layers from Young Shoots of Mature Plants Pinaki Sinha
Tree growth over multiple years
 Tree growth over multiple years All plant growth of originates from meristems or meristematic (capable rapid cell division and differentiation) tissue. Trees are designed to face a problem that annual
Tree growth over multiple years All plant growth of originates from meristems or meristematic (capable rapid cell division and differentiation) tissue. Trees are designed to face a problem that annual
Plant regeneration of Anthurium andreanum cv Rubrun
 Electronic Journal of Biotechnology ISSN: 0717-3458 Vol.7 No.3, Issue of December 15, 2004 2004 by Pontificia Universidad Católica de Valparaíso -- Chile Received June 4, 2004 / Accepted November 9, 2004
Electronic Journal of Biotechnology ISSN: 0717-3458 Vol.7 No.3, Issue of December 15, 2004 2004 by Pontificia Universidad Católica de Valparaíso -- Chile Received June 4, 2004 / Accepted November 9, 2004
MICROPROPAGATION OF JATROPHA CURCAS (L.)
") Indian J. Agric. Res., 43 (4) : 269-273, 2009 AGRICULTURAL RESEARCH COMMUNICATION CENTRE www.arccjournals.com / indianjournals.com MICROPROPAGATION OF JATROPHA CURCAS (L.) B.R. Ranwah, D.K. Gupta and M.A.
Indian J. Agric. Res., 43 (4) : 269-273, 2009 AGRICULTURAL RESEARCH COMMUNICATION CENTRE www.arccjournals.com / indianjournals.com MICROPROPAGATION OF JATROPHA CURCAS (L.) B.R. Ranwah, D.K. Gupta and M.A.
SOMATIC EMBRYOGENESIS AND REGENERATION OF PLANTLET IN SAFFRON, CROCUS SATIVUS L.
 J. Sci. I. R. Iran Vol. 11, No. 3, Summer 2000 SOMATIC EMBRYOGENESIS AND REGENERATION OF PLANTLET IN SAFFRON, CROCUS SATIVUS L. H. Ebrahimzadeh 1*, R. Karamian 2 and M. R. Noori-Daloii 3 1 Department of
J. Sci. I. R. Iran Vol. 11, No. 3, Summer 2000 SOMATIC EMBRYOGENESIS AND REGENERATION OF PLANTLET IN SAFFRON, CROCUS SATIVUS L. H. Ebrahimzadeh 1*, R. Karamian 2 and M. R. Noori-Daloii 3 1 Department of
PlSc 300 Plant Propagation. Final Exam (150 points) Spring 2010
 Spring 2010") 1 PlSc 300 Plant Propagation Final Exam (150 points) Spring 2010 Directions: You have two hours to complete this exam. Try to answer the following questions to the best of your ability. If you have any
1 PlSc 300 Plant Propagation Final Exam (150 points) Spring 2010 Directions: You have two hours to complete this exam. Try to answer the following questions to the best of your ability. If you have any
REGENERATION SYSTEMS FOR PYRAMIDING DISEASE RESISTANCE INTO WALNUT ROOTSTOCKS
 REGENERATION SYSTEMS FOR PYRAMIDING DISEASE RESISTANCE INTO WALNUT ROOTSTOCKS John E. Preece, Ana María Ibáñez, Quyen Tran, Dio Gunawan, Chuck Leslie, David Tricoli, and Abhaya Dandekar ABSTRACT The purpose
REGENERATION SYSTEMS FOR PYRAMIDING DISEASE RESISTANCE INTO WALNUT ROOTSTOCKS John E. Preece, Ana María Ibáñez, Quyen Tran, Dio Gunawan, Chuck Leslie, David Tricoli, and Abhaya Dandekar ABSTRACT The purpose
THE EFFECT OF DIFFERENT PLANT GROWTH REGULATORS ON SHOOT INDUCTION OF Orthosiphon aristatus Boldingh.
 THE EFFECT OF DIFFERENT PLANT GROWTH REGULATORS ON SHOOT INDUCTION OF Orthosiphon aristatus Boldingh. R. Zaidah & M.S. Nazri Biology Department, Faculty of Science, UTM, 81310 UTM Skudai, Johor, Malaysia.
THE EFFECT OF DIFFERENT PLANT GROWTH REGULATORS ON SHOOT INDUCTION OF Orthosiphon aristatus Boldingh. R. Zaidah & M.S. Nazri Biology Department, Faculty of Science, UTM, 81310 UTM Skudai, Johor, Malaysia.
CONSIDERATIONS REGARDING THE EFFECTS OF GROWTH REGULATORS OVER THE IN VITRO MORPHOGENETIC REACTION AT ORIGANUM VULGARE L.
 CRISTEA TINA OANA, FLTICEANU MARCELA, PRISECARU MARIA J. Plant Develop. 15 (2008): 133 138 CONSIDERATIONS REGARDING THE EFFECTS OF GROWTH REGULATORS OVER THE IN VITRO MORPHOGENETIC REACTION AT ORIGANUM
CRISTEA TINA OANA, FLTICEANU MARCELA, PRISECARU MARIA J. Plant Develop. 15 (2008): 133 138 CONSIDERATIONS REGARDING THE EFFECTS OF GROWTH REGULATORS OVER THE IN VITRO MORPHOGENETIC REACTION AT ORIGANUM
Chapter 2 Indirect Organogenesis and histological analysis of organogenic and non-organogenic calli obtained from in vitro
 Chapter 2 Indirect Organogenesis and histological analysis of organogenic and non-organogenic calli obtained from in vitro cultures of Justicia adhatoda L. 2.1. ABSTRACT Leaf, axillary bud and root tip
Chapter 2 Indirect Organogenesis and histological analysis of organogenic and non-organogenic calli obtained from in vitro cultures of Justicia adhatoda L. 2.1. ABSTRACT Leaf, axillary bud and root tip
Application of Plant Hormones
 Application of Plant Hormones Agriculturists all over the world have developed certain unusual methods by which they successfully cultivate the crop plants. It is only in recent year s plant physiologists
Application of Plant Hormones Agriculturists all over the world have developed certain unusual methods by which they successfully cultivate the crop plants. It is only in recent year s plant physiologists
THE CHINCHONA VENEER-GRAFT METHOD OF PROPAGATING SUBTROPICAL FRUIT TREES
 Proc. Fla. State Hort. Soc. 58:176-180. 1945 THE CHINCHONA VENEER-GRAFT METHOD OF PROPAGATING SUBTROPICAL FRUIT TREES William C. Cooper and J. R. Furr United States Department of Agriculture, Orlando Introduction
Proc. Fla. State Hort. Soc. 58:176-180. 1945 THE CHINCHONA VENEER-GRAFT METHOD OF PROPAGATING SUBTROPICAL FRUIT TREES William C. Cooper and J. R. Furr United States Department of Agriculture, Orlando Introduction
BIOLOGY CLASS IX ARTIFICIAL VEGETATIVE PROPAGATION
 BIOLOGY CLASS IX ARTIFICIAL VEGETATIVE PROPAGATION MICROPROPAGATION This a technique of propagating plants by removing tissues from a parent plant and culturing them in a chemical medium. 1. A sterilized
BIOLOGY CLASS IX ARTIFICIAL VEGETATIVE PROPAGATION MICROPROPAGATION This a technique of propagating plants by removing tissues from a parent plant and culturing them in a chemical medium. 1. A sterilized
IN VITRO PROPAGATION OF EGYPTIAN DATE PALM: 11-DIRECT AND INDIRECT SHOOT PROLIFERATION FROM SHOOT-TIP EXPLANTS OF Phoenix dactylifera L. CV. ZAGHLOOL.
 IN VITRO PROPAGATION OF EGYPTIAN DATE PALM: 11-DIRECT AND INDIRECT SHOOT PROLIFERATION FROM SHOOT-TIP EXPLANTS OF Phoenix dactylifera L. CV. ZAGHLOOL. Shawky A. Bekheet and Mahmoud M. Saker Plant Cell
IN VITRO PROPAGATION OF EGYPTIAN DATE PALM: 11-DIRECT AND INDIRECT SHOOT PROLIFERATION FROM SHOOT-TIP EXPLANTS OF Phoenix dactylifera L. CV. ZAGHLOOL. Shawky A. Bekheet and Mahmoud M. Saker Plant Cell
NOTES ON SOME GROWTH CHARACTERISTICS OF MIKANIA CORD AT A (BURM. F.) B.L. ROBINSON*) B.T. MERCADO ABSTRACT INTRODUCTION
 B.L. ROBINSON*) B.T. MERCADO ABSTRACT INTRODUCTION") BIOTROPIA No. 7, 1994: 30-40 NOTES ON SOME GROWTH CHARACTERISTICS OF MIKANIA CORD AT A (BURM. F.) B.L. ROBINSON*) B.T. MERCADO Institute of Biological Sciences, University of the Philippines at Los Banos,
BIOTROPIA No. 7, 1994: 30-40 NOTES ON SOME GROWTH CHARACTERISTICS OF MIKANIA CORD AT A (BURM. F.) B.L. ROBINSON*) B.T. MERCADO Institute of Biological Sciences, University of the Philippines at Los Banos,
4.1. Germplasm collection The state of Tamil Nadu is located in the southern eastern part of Indian
 Results 4.1. Germplasm collection The state of Tamil Nadu is located in the southern eastern part of Indian peninsula 8 o 5` and 13 o 35` North latitude and 76 o 15` and 80 o 20` East longitude. The climate
Results 4.1. Germplasm collection The state of Tamil Nadu is located in the southern eastern part of Indian peninsula 8 o 5` and 13 o 35` North latitude and 76 o 15` and 80 o 20` East longitude. The climate
IN VITRO PROPAGATION OF THE BALKAN ENDEMIC SPECIES VERBASCUM ERIOPHORUM GODR. Abstract. Introduction
 767 Bulgarian Journal of Agricultural Science, 22 (No 5) 2016, 767 771 Agricultural Academy IN VITRO PROPAGATION OF THE BALKAN ENDEMIC SPECIES VERBASCUM ERIOPHORUM GODR. Zh. P. YORDANOVA *, M. A. ROGOVA
767 Bulgarian Journal of Agricultural Science, 22 (No 5) 2016, 767 771 Agricultural Academy IN VITRO PROPAGATION OF THE BALKAN ENDEMIC SPECIES VERBASCUM ERIOPHORUM GODR. Zh. P. YORDANOVA *, M. A. ROGOVA
Physiological and Morphogenetic Studies of Fern Gametophytes and Sporophytes in Aseptic Culture1 IV. Controlled differentiation in leaf callus tissues
 1965 67 Physiological and Morphogenetic Studies of Fern Gametophytes and Sporophytes in Aseptic Culture1 IV. Controlled differentiation in leaf callus tissues Yukio Kato Biological Institute, Faculty of
1965 67 Physiological and Morphogenetic Studies of Fern Gametophytes and Sporophytes in Aseptic Culture1 IV. Controlled differentiation in leaf callus tissues Yukio Kato Biological Institute, Faculty of
Somatic Embryogenesis in Pearl Millet (Pennisetum Glaucum (L) R.Br.) Using Light and Electron Microscopy
 R.Br.) Using Light and Electron Microscopy") IOSR Journal of Agriculture and Veterinary Science (IOSR-JAVS) e-issn: 2319-2380, p-issn: 2319-2372. Volume 5, Issue 1 (Sep.- Oct. 2013), PP 01-09 Somatic Embryogenesis in Pearl Millet (Pennisetum Glaucum
IOSR Journal of Agriculture and Veterinary Science (IOSR-JAVS) e-issn: 2319-2380, p-issn: 2319-2372. Volume 5, Issue 1 (Sep.- Oct. 2013), PP 01-09 Somatic Embryogenesis in Pearl Millet (Pennisetum Glaucum
HORMONE ROOT-SOAK CAN INCREASE INITIAL GROWTH OF PLANTED HARD- WOOD STOCK 1
 Winter 1980/29 HORMONE ROOT-SOAK CAN INCREASE INITIAL GROWTH OF PLANTED HARD- WOOD STOCK 1 R. C. Hartwig and M. M. Larson Graduate Student and Professor, Department of Forestry, Ohio Agricultural Research
Winter 1980/29 HORMONE ROOT-SOAK CAN INCREASE INITIAL GROWTH OF PLANTED HARD- WOOD STOCK 1 R. C. Hartwig and M. M. Larson Graduate Student and Professor, Department of Forestry, Ohio Agricultural Research
MICROPHOPAGATION OF DATE PALM (PHOENIX TISSUE CULTURE TECHNIQUE
 Annals of Arid Zone 28 (1&2): 137-141, 1989 MICROPHOPAGATION OF DATE PALM (PHOENIX DACTYLIFERA L.) CV KHADRA WY USING TISSUE CULTURE TECHNIQUE N.L. KACKAR, K.R. SOLANKI AND S.P. JOSHI Central Arid Zone
Annals of Arid Zone 28 (1&2): 137-141, 1989 MICROPHOPAGATION OF DATE PALM (PHOENIX DACTYLIFERA L.) CV KHADRA WY USING TISSUE CULTURE TECHNIQUE N.L. KACKAR, K.R. SOLANKI AND S.P. JOSHI Central Arid Zone
Source Published by. The Singapore Copyright Act applies to the use of this document. Archived with permission from the copyright owner.
 Title Author(s) Source Published by Plantlet production of Swietenia macrophylla King through tissue culture A. N. Rao and S. K. Lee Gardens Bulletin Singapore, 41, 11-18 National Parks Board, Singapore
Title Author(s) Source Published by Plantlet production of Swietenia macrophylla King through tissue culture A. N. Rao and S. K. Lee Gardens Bulletin Singapore, 41, 11-18 National Parks Board, Singapore
Agrobacterium Mediated Tobacco Transformation By: Patrick Kudyba Section: 002 April 15, 2015
 Agrobacterium Mediated Tobacco Transformation By: Patrick Kudyba Section: 002 April 15, 2015 Introduction Transformation is an important tool to plant biotechnologists because it is an easy way to insert
Agrobacterium Mediated Tobacco Transformation By: Patrick Kudyba Section: 002 April 15, 2015 Introduction Transformation is an important tool to plant biotechnologists because it is an easy way to insert
FERN MULTIPLICATION KIT
 FERN MULTIPLICATION KIT Product No. F34 India Contact 1 2 3 6 I 7...7 Product Number Kit Components Product Description 2 1 EA Box 1 Instruction Manual 1 C913/C21-10ea Culture Containers 1 F91-1ea Forceps,
FERN MULTIPLICATION KIT Product No. F34 India Contact 1 2 3 6 I 7...7 Product Number Kit Components Product Description 2 1 EA Box 1 Instruction Manual 1 C913/C21-10ea Culture Containers 1 F91-1ea Forceps,
International Journal of Pharma and Bio Sciences RAPID IN VITRO PROPAGATION TECHNIQUE FOR SUGARCANE VARIETY 018
 International Journal of Pharma and Bio Sciences RESEARCH ARTICLE BIOTECHNOLOGY RAPID IN VITRO PROPAGATION TECHNIQUE FOR SUGARCANE VARIETY 018 SATPAL SINGH BISHT*, AJIT KUMAR ROUTRAY & ROJITA MISHRA Department
International Journal of Pharma and Bio Sciences RESEARCH ARTICLE BIOTECHNOLOGY RAPID IN VITRO PROPAGATION TECHNIQUE FOR SUGARCANE VARIETY 018 SATPAL SINGH BISHT*, AJIT KUMAR ROUTRAY & ROJITA MISHRA Department
Regeneration of Plantlets Through PLB (Protocorm-Like Body) Formation in Phalaenopsis Join Angle X Sogo Musadian
 Formation in Phalaenopsis Join Angle X Sogo Musadian") 204 J. Math. Fund. Sci., Vol. 48, No. 3, 2016, 204-212 Regeneration of Plantlets Through PLB (Protocorm-Like Body) Formation in Phalaenopsis Join Angle X Sogo Musadian Dini Meilasari & Iriawati School
204 J. Math. Fund. Sci., Vol. 48, No. 3, 2016, 204-212 Regeneration of Plantlets Through PLB (Protocorm-Like Body) Formation in Phalaenopsis Join Angle X Sogo Musadian Dini Meilasari & Iriawati School